|
Intervention Theory Essentials |
|
 |
Intervention Theory Essentials
By Lloyd Pye
A Basic Guide to the Intervention Theory of Origins
— of the Universe, of Life, of Hominoids, and of Humans.
Version 10.20
© Lloyd Pye, 2011
This eBook is 350 pages in a large font for ease of reading on computers and handhelds. It has 230 photos and illustrations. It contains 40,000 words. Most people read at 200 to 250 words per minute, so this will require about 3 hours.
Those 3 hours will rock your worldview.
“When you seek a path to any new truth, you must expect to find it blocked by ‘expert opinion.’” – Albert Guérard
“An emotional plague afflicts people whose belief systems are so rigid they ignore relevant facts and become enraged if anyone challenges their beliefs.” – Wilhelm Reich
“For every PhD., there is an equal and opposite PhD.” – Gibson’s Law
“Anyone can pilot an improbability, but it takes a special touch to fly and safely land an absurdity.” – Kurt Kuzba
“A genius is someone who aims at a target no one else can see — and hits it!” – Anthony Quinn
“The main purpose of science is to investigate the unexplained, not to explain the uninvestigated.” Dr. – Stephen Rorke
“The supreme arrogance of religious thinking is that a carbon-based bag of mostly water on a speck of iron- silicate dust around a boring dwarf star in a minor galaxy in an unfashionable suburb of (our) supercluster would look up at the sky and declare: “It was all made so that I could exist!”” – Peter Walker
In 1905, a 25-year-old patent clerk named Albert Einstein demolished the 200-year-old certainty that Isaac Newton knew all there was to know about basic physics.
- In a technical paper only a few pages long, Einstein sent a huge part of his current "reality" to history's dustbin, where it found good company with thousands of other discards large and small.
- In 1905, though, Newton's discard was about as large as the bin would hold.
Now another grand old "certainty" hovers over history's dustbin, and it seems only a matter of time before some new Einstein writes the few (or many) pages that will bring it down and relegate it to history.
- And, as was the case in 1905, every "expert" in the world laughs heartily at any suggestion that their certainty could be struck down.
- Yet if facts are any yardstick — which should always be the case, but frequently isn't
— Charles Darwin's theory of evolution by natural selection is moving towards extinction.
Please note this: not everyone who challenges evolution is automatically a Creationist. Darwinists love to tar all opponents with that brush because so much of Creationist dogma is absurd. Creationists mulishly exclude themselves from serious consideration by refusing to give up fatally flawed parts of their argument, such as the literal interpretation of "six days of creation". Of course, some have tried to take a more reasonable stance, but those few can't be heard over the ranting of the many who refuse.
Recently a new group has entered the fray, much better educated than typical Creationists. This group has devised a theory called "Intelligent Design", which has a wealth of scientifically established facts on its side. The ID-ers, though, give away their Creationist roots by insisting that because life at its most basic level is so incredibly and irreducibly complex, it could never have simply "come into being" as Darwinists insist.
Actually, the "life somehow assembled itself out of organic molecules" dogma is every bit as absurd as the "everything was created in six days" dogma, which the ID-ers understand and exploit. But they also suggest that everything came into existence at the hands of God (by whatever name) or "by means of outside intervention", which makes clear how they're betting. "Outside intervention" is a transparent euphemism for "You Know What" (with apologies to J. K. Rowling) In Rowling's "Harry Potter" books, the arch villain is so despicable and dreadful, his name should not even be uttered; thus he is referred to as "You Know Who".
Similarly, the very idea that humans might have been created by extraterrestrials is so despicable and dreadful to mainstream science and religion that no mention of it should be uttered; thus the author refers to it as "You Know What". To Darwinists, Creationists and ID-ers alike, creation at the hands of You Know What is the most absurd suggestion of all. Yet it can be shown that You Know What has the widest array of facts on its side and has the best chance of being proved correct in the end.
Virtually every scientist worth their doctorate will insist that somehow, some way, a form of evolution is at the heart of all life forms and processes on Earth. By "evolution", they mean the entire panoply of possible interpretations that might explain how, over vast stretches of time, simple organisms can and do transform themselves into more complex organisms. That broad definition gives science as a whole a great deal of room to bob and weave its way towards the truth about evolution, which ostensibly is its goal. However, among individual scientists that same broadness of coverage means nobody has a "lock" on the truth, which opens them up to a withering array of internecine squabbles.
In Darwin's case, those squabbles were initially muted. Rightly or wrongly, his theory served a much higher purpose than merely challenging the way science thought about life's processes. It provided something every scientist desperately needed: a strong counter to the intellectual nonsense pouring from pulpits in every church, synagogue and mosque in the world.
Since well before Charles Darwin was born, men of science knew full well that God did not create the Earth or anything else in the universe in six literal days. But to assert that publicly invited the same kind of censure that erupts today onto anyone who dares to challenge evolution openly. Dogma is dogma in any generation.
Darwin's honeymoon with his scientific peers was relatively brief. It lasted only as long as they needed to understand that all he had really provided was the outline of a forest of an idea, one that only in broad terms seemed to account for life's stunningly wide array. His forest lacked enough verifiable trees.
Even so, once the overarching concept was crystallized as "natural selection", the term "survival of the fittest" was coined to explain it to laymen. When the majority of the public became convinced that evolution was a legitimate alternative to Creationism, the scientific gloves came off. In-fighting became widespread regarding the trees that made up Darwin's forest.
Over time, scientists parsed Darwin's original forest into more different trees than he could ever have imagined. That parsing has been wide and deep, and it has taken down countless trees at the hands of scientists themselves. But despite such thinning, the forest remains upright and intact. Somehow, some way, there is a completely natural force at work governing all aspects of the flow and change of life on Earth. That is the scientific mantra, which is chanted religiously to counter every Creationist — and now Intelligent Design — challenge to one or more of the rotten trees that frequently become obvious.
Even Darwin realized the data of his era did not provide clear-cut evidence that his theory was correct. Especially troubling was the absence of "transitional species" in the fossil record. Those were needed to prove that, over vast amounts of time, species did in fact gradually transform into other, "higher" species.
So right out of the chute, the theory of evolution was on the defensive regarding one of its cornerstones, and more than 140 years later there are still no clear-cut transitional species apparent in the fossil record.
Because this is the most vulnerable part of Darwin's theory, Creationists attack it relentlessly, which has forced scientists periodically to put forth a series of candidates to try to take the heat off. Unfortunately for them, in every case those "missing links" have been shown to be outright fakes and frauds. An excellent account is found in Icons of Evolution by Jonathan Wells (Regnery, 2000). But scientists are not deterred by such exposure of their shenanigans. They feel justified because, they insist, not enough time has passed for them to find what they need in a grossly incomplete fossil record.
The truth is that some lengthy fossil timelines are missing, but many more are well accounted for. Those have been thoroughly examined in the past 140-plus years, to no avail. In any other occupation, a 140-year-long trek up a blind alley would indicate a wrong approach has been taken. But not to scientists.
They blithely continue forward, convinced of the absolute rightness of their mission and confident their fabled missing link will be found beneath the next overturned rock. Sooner or later, they believe, one of their members will uncover it, so they all work in harmonious concert towards that common goal. Individually, though, it's every man and woman for themselves.
To Daniel Shechtman, Ph.D. Winner of the 2011 Nobel Prize for Chemistry.
In 1982, Dr. Shechtman created a crystal with atoms that formed a five-sided pattern that did not repeat itself. He defied a “received wisdom” in chemistry that they must create “repetitious” patterns such as triangles, squares or hexagons.
Shechtman’s peers ridiculed his discovery of what has come to be known as quasi-crystals. Prior Nobel laureate Linus Pauling castigated him by insisting: “There is no such thing as quasi-crystals, only quasi-scientists.”
After years of battling the dogma of ignorance, Shechtman’s efforts finally forced his peers to reconsider the fundamental nature of matter.
This is how it always is with science. Someone discovers a new concept, and first it is ignored, then it is ridiculed, and finally it is self-evident
This eBook is designed to explain the essential aspects of Intervention Theory that anyone new to it, or interested in learning more about it, will need to know. It is grounded in solid, reliable academic research, though it is not footnoted or annotated because there is no point in doing so.
Statements made by alternative researchers like me are automatically contradicted by scientists insisting we are not simply wrong, but stupidly wrong. They further insist we have no right to challenge their cherished beliefs because our only “credentials” are an unwarranted faith in our ability to discern truth from nonsense.
[This doesn’t refer to all scientists. Some still willingly risk reputation and security to explore topics that defy dogma. However, they are few.]
Every point I discuss is supported by facts in available research, but I am often criticized by skeptics. Why? Because the issues I discuss are long-lived sacred cows to mainstream sciences.
They protect their herd with admirable tenacity, as I would if I had somehow joined their ranks.
Looking back, I’m delighted I managed to find a different herd to look after, because I’m sure those areas of study will lead to our true future.
Also, this eBook contains a small amount of repetition. The best way to learn anything, to make it stick in memory like velcro, is to use a technique known as Programmed Learning.
Years ago, school workbooks had sentences containing blank segments to be filled in by students. They were intermittently repetitive, which subtly enhanced memory. Workbooks are no longer used as much, but intermittent repetition remains a useful learning tool.
This eBook’s purpose is to introduce readers to highly controversial concepts they should easily absorb and, more importantly, recall with some detail for later discussion and, if needed, debate with friends and family not yet familiar with the basic evidence supporting Intervention Theory.
Plants and animals evolve, eh? Alright, how do they evolve?
- By gradual but constant changes, influenced by adaptive pressures in their environment that cause physical modifications to persist if they are advantageous.
Can you specify the kind of gradual change you're referring to?
- In any population of plants or animals, over time, random genetic mutations will occur. Most will be detrimental, some will have a neutral effect and some will confer a selective advantage, however small or seemingly inconsequential it might appear.
Really? But wouldn't the overall population have a gene pool deep enough to absorb and dilute even a large change? Wouldn't a small change rapidly disappear?
- Well, yes, it probably would. But not in an isolated segment of the overall population. An isolated group would have a much shallower gene pool, so positive mutations would stand a much better chance of establishing a permanent place in it.
Really? What if that positive mutation gets established in the isolated group, then somehow the isolated group gets back together with the main population? Poof! The mutation will be absorbed and disappear.
- Well, maybe. So let's make sure the isolated population can't get back with the main group until crossbreeding is no longer possible.
How would you do that?
- Put a mountain range between them, something impossible to cross. If it's impossible to cross, how did the isolated group get there in the first place?
If you're asking me just how isolated is isolated, let me ask you one. What kind of mutations were you talking about being absorbed?
- Small, absolutely random changes in base pairs at the gene level.
Really? Why not at the chromosome level? Wouldn't change at the base pair level be entirely too small to create any significant change? Wouldn't a mutation almost have to be at the chromosome level to be noticeable?
- Who says? Change at that level would probably be too much, something the organism couldn't tolerate. Maybe we're putting too much emphasis on mutations.
Right! What about environmental pressures? What if a species suddenly found itself having to survive in a significantly changed environment?
- One where its members must adapt to the new circumstances or die out?
Exactly! How would they adapt? Could they just will themselves to grow thicker fur or stronger muscles or larger size?
- That sounds like mutations have to play a part.
Mutations, eh? All right, how do they play a part?
This game of intellectual thrust and parry goes on constantly at levels of minutiae that boggle an average mind.
- Traditional Darwinists are one-upped by neo-Darwinists at every turn.
- Quantum evolutionists refashion the work of those who support the theory of peripheral isolates.
- Mathematicians model mutation rates and selective forces, which biologists do not trust.
- Geneticists have little use for paleontologists, who return the favor in spades (pun intended).
- Cytogenetics labours to find a niche alongside genetics proper.
- Population geneticists utilize mathematical models that challenge paleontologists and systematists.
- Sociobiologists and evolutionary psychologists struggle to make room for their ideas.
- All perform a cerebral dance of elegant form and exquisite symmetry.
Their dance is, ironically, evolution writ large throughout science as a process. New bits of data are put forth to a peer group.
The new data are discussed, written about, criticized, written about again, criticized some more. This is gradualism at work, shaping, reshaping and reshaping again if necessary until the new data can comfortably fit into the current paradigm in any field, whatever it is. This is necessary to make it conform as closely as possible to every concerned scientist's current way of thinking. To do it any other way is to invite prompt rejection under a fusillade of withering criticism.
This system of excruciating "peer review" is how independent thinkers among scientists have always been kept in line. Darwin was an outsider until he barged into the club by sheer, overpowering brilliance. Patent clerk Einstein did the same. On the other hand, Alfred Wegener was the German meteorologist who figured out plate tectonics in 1915. Because he dared to bruise the egos of "authorities" outside his own field, he saw his brilliant discovery buried under spiteful criticism that held it down for 50 years. Every scientist in the game knows how it is played, and very few dare to challenge its rules.
The restrictions on scientists are severe, but for a very good reason. They work at the leading edges of knowledge, from where the view can be anything from confusing to downright terrifying. Among those who study the processes of life on Earth, they must cope with the knowledge that a surprising number of species have no business being here. In some cases, they can't even be here. Yet they are, for better or worse, and those worst-case examples must be hidden or at least obscured from the general public. But no matter how often facts are twisted, data are concealed or reality is denied, the truth is out there.
 |
Creationists of all kinds … “God did it!”
Darwinists of all stripes … “It just happened — poof! — like magic!”
Outside Intervention … “They did it!”
This is how the Bible explains it …
In the beginning was the Word, and the Word was with God, and the Word was God.
Yet, to account for all the magnificence that has come into existence in our universe since then, that Word must have been on the scale of:
S.u.p.e.r.c.a.l.i.f.r.a.g.i.l.i.s.t.i.c.e.x.p.e.a.l.i.d.o.c.i.o.u.s
Creationists of all kinds, which includes the smarter, more reasonable, and typically well-credentialed Intelligent Design proponents, insist the only answer to questions about the origins of life or humans is: “God did it!”
Darwinists of all stripes insist with equal zeal that their pet theory of evolution best explains how life originated. Their 150-year-old dogma asserts: “It just happened — poof! — like magic!”
After the magic moment when life kick-started itself into existence, it initiated a self-contained and imperceptibly slow-but-steady growth into ever more complex forms, until those reached the apex of an arduous climb — humanity.
In contrast, Interventionists like me anchor our search for origins on evidence rather than faith, on logic rather than magic. We don’t think that God did it, or that life spontaneously generated.
For us, evidence and logic point to the same “outside intervention” Intelligent Designers see. However, where they feel the only outside source of intervention must be God (whom they are careful to not mention by name), we suggest another, bolder explanation: “They did it!”
Who are “They”?
The currently favored term is Aliens — non-human, non-Earth-based entities.
Of course, aliens raises the hackles and blood pressure of science, government, and religion, so to calm them I will later provide a different, less threatening term. That new term describes entities who have created and distributed, then overseen and managed, life’s myriad forms.
Mainstream scientists say extraordinary claims require extraordinary evidence. [In this eBook I will forego the common jibe, Lamestream, out of respect for its ideals rather than its practices.]
Clearly, the Intervention Theory makes several super-extraordinary claims, so we need a great deal of extraordinary evidence. Do we have it?
This eBook is meant to answer those questions with facts, data and evidence that in the court of public opinion should qualify as extraordinary.
That human life emerged from primitive haircovered hominoids (upright walking apes) after human-like entities (aliens or gods, with a small “g”) intervened genetically (with test tubes) to create a new hybrid being (humans) with genes from themselves and the primitive hominoids.
 |
Cosmologists and astrophysicists (collectively I’ll call them cosmologists) believe the universe began with a colossal Big Bang. The amusing poster here summarizes their grandiose theory.
Obviously, that theory needs some work, but the cosmologists are stuck with it because it is based on theories and “received wisdom” they see as unassailable, with all of it founded on the idea that gravity is the inherent physical force that binds together everything in the universe.
This received wisdom came down to them from Isaac Newton, who in 1687 first grappled with trying to figure out gravity and how it functions not only on the Earth, but in our solar system as a whole, and in the infinite universe beyond.
In the early 1900s, Albert Einstein added to the gravitational riddle cosmology confronted when he wove electromagnetism into the mix. But no matter how hard cosmologists tried, they nor Einstein could make the gravity math add up.
 |
It was clear that something was missing in the gravity-based formulas. To make them work, they had to mimic a classic S. Harris cartoon:
Gravity-based math was woefully inadequate to explain the real forces at work in the universe. How to solve this conundrum? Easy …. cheat! Cosmologists realized they could add abstract “fudge factors” to provide the miracles their formulas required — as many as they needed!
 |
To make gravity-based math work as it should, cartoonish concoctions were added to the faulty equations. Dark Matter, Dark Energy, Black Holes and, yes, even the Big Bang itself came from their tinkering with reality to make it fit what they needed it to be rather than what it is.
Calling these enormities “fudge factors” is an exercise in chutzpa. Gravity-based math says 96%* of the universe is missing! Not there! [*Different sources give different percentages.]
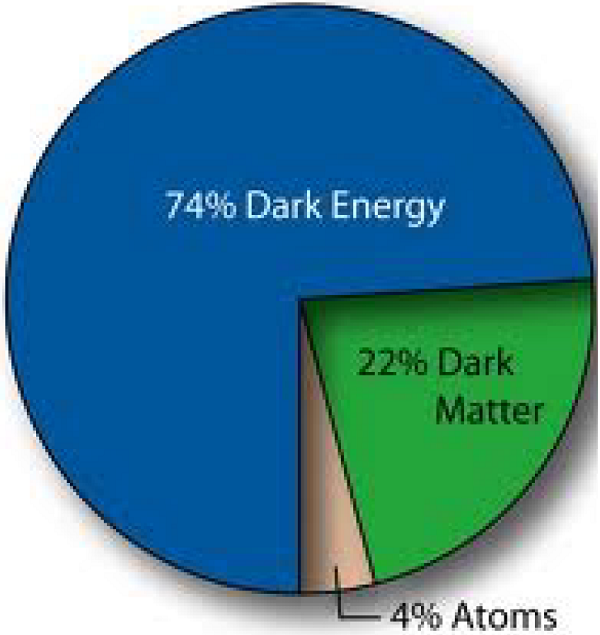 |
Dark Energy, Dark Matter, and Black Holes had to be fabricated to supplement a Big Bang and obscure gargantuan gaps in cosmic reality.
What is the real problem here? What are they so desperate to avoid having to confront? The bottom line is, indeed, math … the math that simply won’t add up for them the way it must for Newton and Einstein to remain relevant.
 |
Imagine giving up Newton and Einstein. Doing that would decimate everything cosmology was structured around and built upon. It would force every “expert” back to classrooms to start over. It would mean rewriting all of their textbooks!
Who in their right mind would want a disaster like that to happen during their watch? So, they protect gravity with religious zeal, even though it is clear to anyone who studies the problem to any depth that they are riding a lame horse.
Here is the number that lames their horse: 1039 (read: 10 to the power of 39). It is 10 followed by 39 zeroes. It is a mind-boggling number. So, what is it? What does it represent? It expresses the power of electricity compared to gravity.
As a force, electricity is 39 orders of magnitude stronger than gravity!!! That is one thousand billion billion billion billion times stronger!
Cosmologists should have ditched gravity as soon as they understood that imposing number. Unfortunately, they made the decision to keep doing what they had always done rather than forging into unfamiliar territories where their ignorance would be obvious and embarrassing as they learned that new turf. They decided to stay hidden behind gravity’s deceiving skirts.
Gravity can’t exist without an electromagnetic force. It is dependent on the electrified plasma that thinly permeates the universe. Plasma is the cohesive agent cosmologists claim for gravity.
Gravity only has impact as a force when it is contained in enormous celestial bodies (suns, planets, moons). At the granular level, where gravity supposedly draws particles together in the vacuum of space to create everything in the universe, it has literally no power! It is useless!
This explains why massive cheating is needed to make the gravity math work. But, in contrast, if the immense power of electricity is plugged into the same formulas, the math works without fudge factors. The universe’s “missing” 96% becomes mathematically viable, and reality is served rather than cosmology’s fragile egos.
 |
For those who think I might be off base here, let me refer you to the same sources I used to locate this paradigm-busting information. Two marvelous websites that support what is called The Electric Universe Theory are found here:
- www.Thunderbolts.info
- www.Holoscience.com
- Both are excellent!
I have started this eBook with cosmology and astrophysics because they are the sciences most concerned with how the universe began, and because their dogmatism illustrates why I keep insisting Everything You Know Is Wrong. This is one of numerous examples I will provide to support Intervention Theory as we move on.
The sad truth is that in every field of science, Young Turks have to serve their leaders when those leaders are the oldest, crustiest, and most conservative members of the field. Then, when those old “defenders of the faith” die out, what were once Young Turks take over for them and are forced to defend the same bankrupt faith.
 |
Cosmologists tell us our solar system began as a cloud of dust and gas swirling as a disk that separated into the sun, planets, moons, comets, and asteroids. This scenario could be accurate.
However, as previously noted, gravity could not cause any granular particles to aggregate in the vacuum of space. That had to be caused by the electromagnetism generated by the electrified plasma that exists throughout the universe.
Mainstream geologists have concluded that the solar system began to coalesce into its various planets and moons at around 4.5 billion years ago (bya). Agreement among experts does not mean their opinions are reliable, but in this case I accept their conclusion as a reasonable guess.
 |
Another mainstream consensus is that during the proto-Earth’s first half-billion years it was a cauldron of seething lava relentlessly impacted by meteors and asteroids tearing through space.
This hellish period is called the Early Heavy Bombardment (EHB), from 4.5 bya to 4.1 bya, followed by the even more destructive impacts of the Late Heavy Bombardment (LHB), 4.1 to 3.8 bya, and notable for the smaller craters it left on the proto-Moon and the proto-Earth.
[Large Moon craters may not be from impacts. The Moon must be a topic in another eBook.]
Despite the EHB and LHB, in those 0.7 billion years proto-Earth steadily cooled and solidified in the sub-zero temperatures of space. The first rocks seem to have formed at around 3.8 bya.
Science teaches that those rocks were in small masses of land that gradually grew as volcanoes spewed more lava. With eruptions came steam that condensed into the first puddles of water.
[Heated debate centers on why so much water is now on Earth. This too is for another eBook.]
We are told that at 2 bya, surface cooling was complete. Dry land that resembled today’s lava fields was widespread, as was abundant water in collections that ranged from shallow pools to ponds to lakes to seas. Earth had become Earth.
 |
Now, what about the origin of life? When did it occur? How did it occur? The mainstream gives two options and rejects a third:
- They accept as a possibility Undirected Panspermia. This is the idea that life “drifts” across space carried on asteroids or meteors that crashed into Earth.
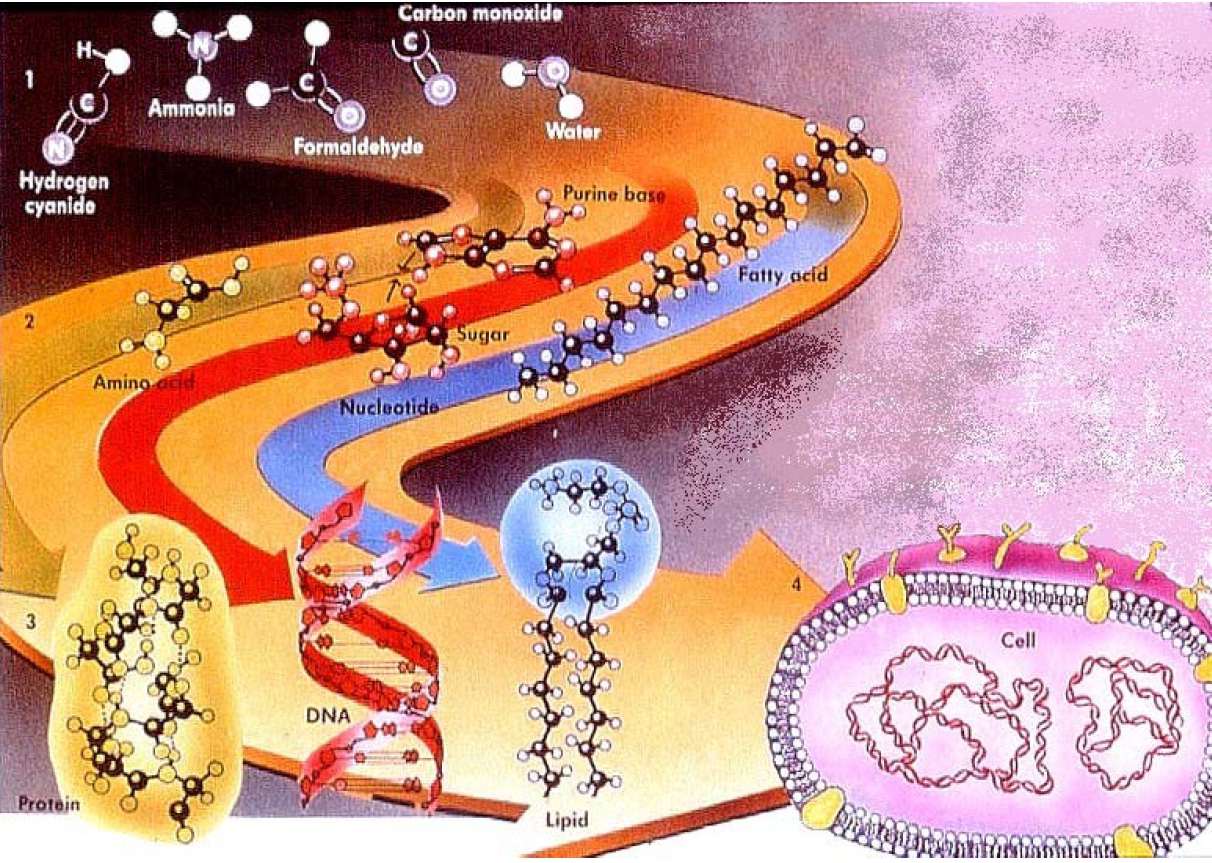
- They support the idea that life originates by Spontaneous Generation. This idea is that life was created by sheer chance when a lightning bolt struck a “warm pond” (a term coined by Charles Darwin) filled with what came to be called “primordial soup,” a form of witches’ brew containing the “building blocks” of life.
The Primordial Soup Theory is shown at right. Reality extends only as far as the top part of the image. Molecules like the five shown exist now and almost certainly existed in the distant past. Then, according to scientific imaginings, they moved onto a magic yellow brick road to life.
Experts insist that somehow, someway, those original molecules managed to spontaneously reassemble into ever more complex molecules that somehow, someway, formed themselves into the single cells of the earliest life forms.
 |
The fact that spontaneous self-assembly into complex molecules does not happen now, nor can it be forced to happen in coherent patterns in the most sophisticated laboratories on Earth, should be an indication that this theory, like the gravity-based one of cosmologists, needs work.
British astrophysicist Fred Hoyle summed up the problem neatly when he said the likelihood of a lightning bolt striking water to assemble simple molecules into a life form was equal to the likelihood of a tornado sweeping through a junkyard and correctly assembling a jetliner.
 |
The third option, which the mainstream rejects, is Directed Panspermia, which is similar to undirected Panspermia, except that the first life forms were guided here by higher intelligence.
Thus, the mainstream hangs its hat on a miracle worthy of God when a lightning bolt struck the “primordial soup” to forge basic molecules into simple life — or on a similarly unlikely miracle that simple life forms came here on meteors.
The lightning-bolt-hits-primordial-soup theory is still taught around the world, but not because mainstream scientists regard it seriously. They know as well as anyone how absurd it sounds.
Unfortunately, they have no choice but to fake conviction about it because they have no other theory to take its place. Some few scientists do admit they have no plausible idea, but most of them insist on riding the same old lame horse.
Like the equations of the cosmologists, which required “miracles” to make sense, the same is true for biologists. The same kinds of miracles are fudge-factored in to explain life, when they have answers that are as easy to grasp as the difference between gravity and electricity.
In the same way cosmologists stick with the mistake of gravity, biology’s mistake, the core of its ossified dogma, is that every aspect of life on Earth must be accounted for in a “natural” manner, in strictly terrestrial terms. To explain life by using “outside” factors is unacceptable, so alternate ideas are automatically discounted.
Despite automatic rejection, alternative ideas for the origin of life make a much stronger case than biological dogma. Such ideas also hew far more closely to the actual facts of life than do the fantasies created by imaginative scientists.
Life did not start by accident when lightning struck a warm pond of primordial soup. Its start was so unlikely, experts can’t begin to explain it rationally, so rather than try, they obscure it.
 |
How could simple molecules like ammonia, carbon monoxide, formaldehyde, hydrogen cyanide, and water, floating loose in Earth’s environment, turn into nucleic acids directing proteins and lipids to make cell membranes?
That didn’t happen because it can’t happen, so there must be another answer … and there is! It turns out life suddenly appears on Earth — at a highly improbable time, in a highly improbable form, in a highly improbable manner. Let’s set aside its improbability to ask how it appeared.
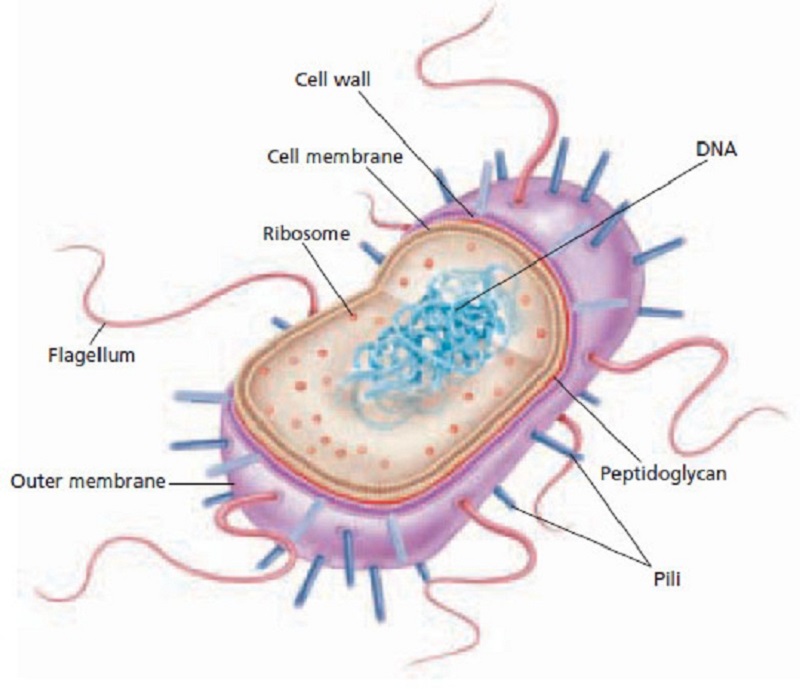
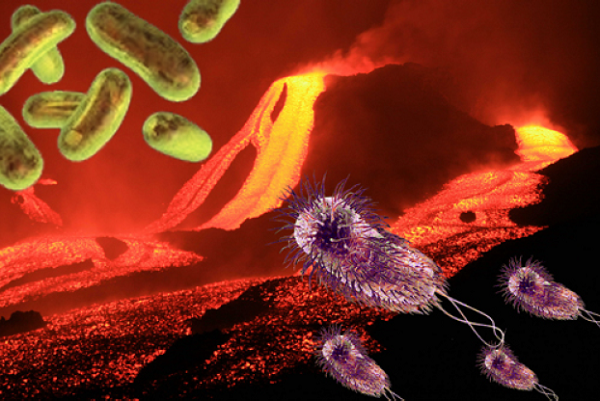
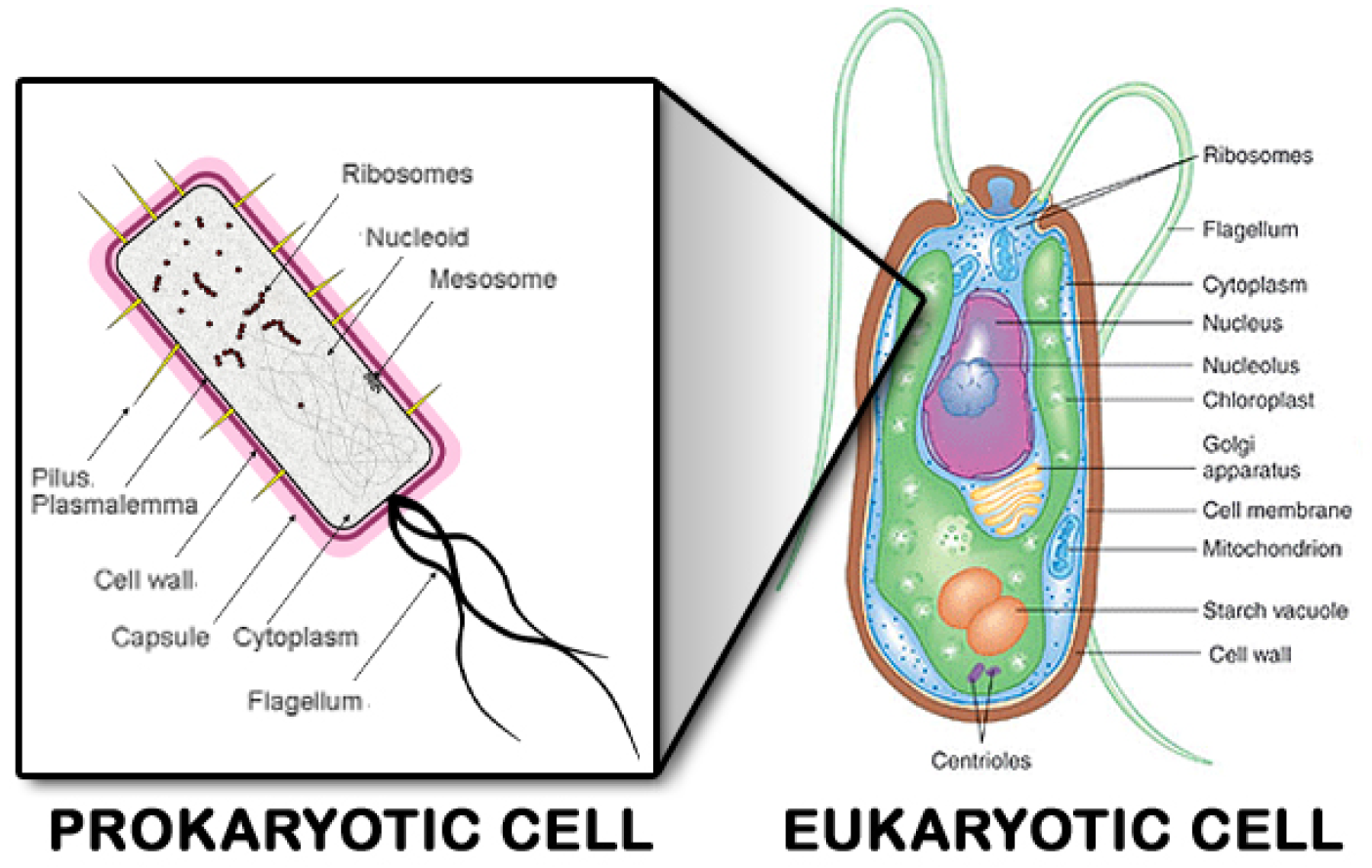
The image to the right is a prokaryote. It represents millions of types that currently exist, and all are complex one-celled bacteria with DNA strands floating free in the cell’s cytoplasm rather than being encased in a nucleus. That comes later.
Prokaryotes (pro-carry-oats) today are adapted to current conditions, but many types survive in the extreme environments of the earliest Earth. They are anaerobic (living without oxygen).
They thrive in salt, sulphur, methane, boiling heat, freezing cold, or under high pressures at great depths. In doing so, many — but not all — of their metabolic processes produce oxygen.
Given their extreme durability and capacity to pump oxygen into the environment, they would be perfect “seeds” to establish life on any protoplanet. If you wanted to spread life across the galaxy, prokaryotes would be the ideal tool.
 |
Imagine certain entities of indeterminate origin decided to ensure life would be given a chance to take root and thrive on any of the 50 million planets in our galaxy where it might flourish. How could those entities be sure it happened?
How about loading an enormous spaceship with the most durable prokaryotic bacteria, and then setting out to visit all of the new solar systems forming across our galaxy? Disregard the issue of the great distances involved. These entities learned long ago how to cruise the galaxy as easily as we drive around our neighborhoods.
Okay, a freshly coalescing solar system looms ahead. What do we do? Wait a billion or more years for its planets and moons to cool? Naaah! Why bother? We have prokaryotes on board!
They can live anywhere, so let’s go ahead and dump them onto the scalding lava beds spewing from the active volcanoes, and into the seething
collections of condensed steam, both of which will later turn into dry land and pools of water.
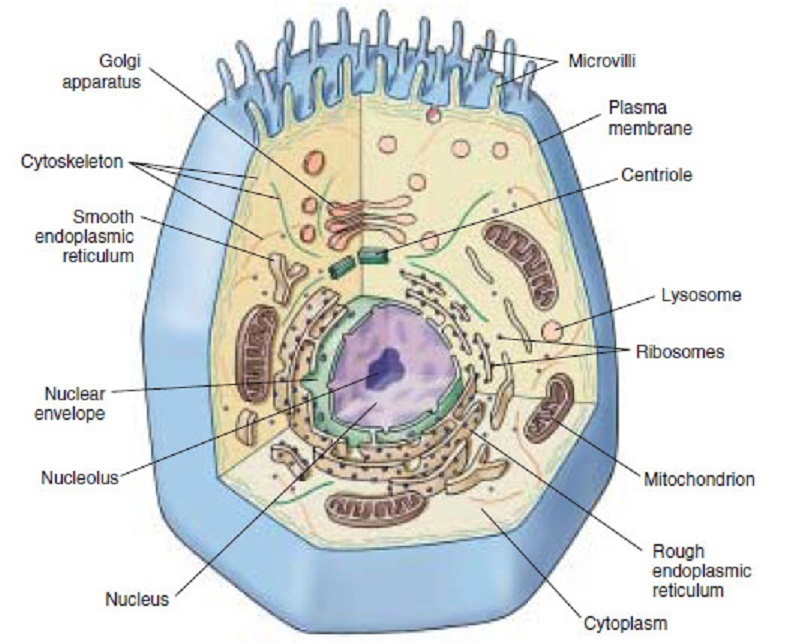 |
Why waste one or two billion years waiting for cooling when the metabolism of indestructible anaerobic prokaryotes can put oxygen into the air and water of any congealing planets? That allows those planets to move to the next phase of complex life: eukaryotes (you-carry-oats).
Eukaryotes (pictured at right) are much larger and vastly more complex than prokaryotes. The simplest types remain single-celled bacteria, but they have a radically different internal design that encases their DNA in a central nucleus. Also, some are anaerobic oxygen producers, while others are aerobic, using oxygen to produce other gases.
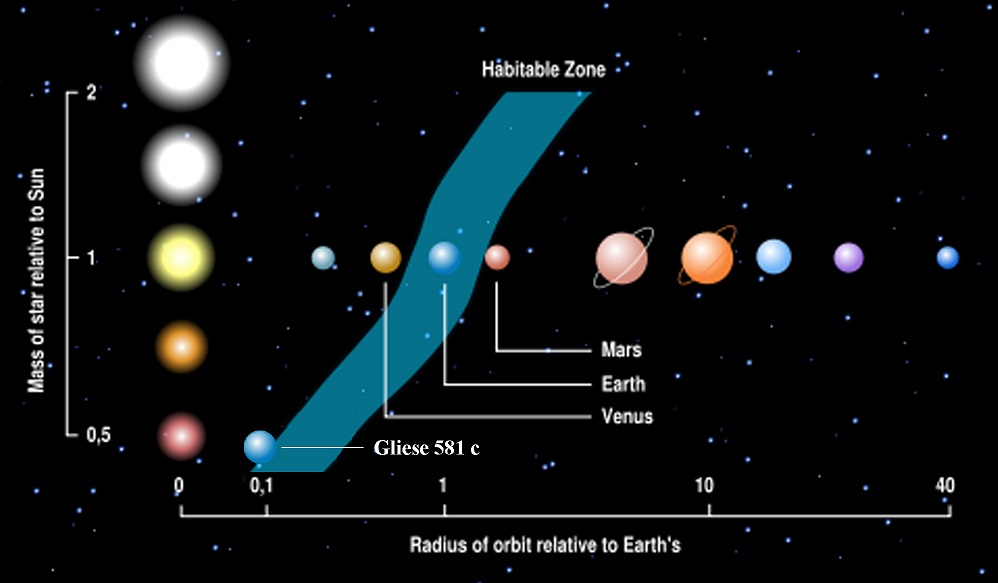 |
The much larger eukaryotes would give a huge advantage to our Intragalactic Terraformers, because eukaryote metabolism puts much more oxygen into the air and water of any planet in what we consider The Goldilocks Zone — the area not too hot, not too cold, but “just right.”
We assume the Goldilocks Zone of a star of any size (blue stripe) can support the life cycles of multi-cellular life forms, up to highly complex forms that contain species and subspecies.
Right now, Earth is the only planet we can be certain fully utilizes its place in the Goldilocks Zone around our Sun, but astronomers find new stars with planets in their Goldilocks Zones on a regular basis. That doesn’t mean they support complex life, because we don’t fully understand the vagaries of life beyond Earth’s boundaries.
 |
Many experts insist life is too complex to have formed anywhere except on Earth. Others feel its exceeding complexity ensures that if it exists elsewhere, it will have a uniform genetic code. To presume it spontaneously assembles around different templates seems highly improbable.
With Earthly life so complex, it seems safe to presume the same structure exists everywhere. Likewise, because Earth contains millions of different life forms, the famous bar scene in the movie Star Wars may be entirely reasonable.
 |
As our solar system formed, any Intragalactic Terraformers passing by could not have known which planets would end up in the zone of life.
Also, the Goldilocks Zone is a construct of our current understanding of how life works on our Sun and Earth. A larger area around other suns might support complex varieties of life we can’t begin to imagine. The depth of our ignorance about these matters is certain to be profound.
Let’s assume that everywhere life exists — on Earth and beyond — it is every bit as resilient and prolific as here. Whether that is true or not, we do know that myriad varieties are here, and we know when the first prokaryotes appeared.
Despite fanciful dogmas experts preach to the uninformed, when prokaryotes first appeared, Earth was as depicted above, as it would have been if our imagined Intragalactic Terraformers were actually at work. And if they were, they provide a much more plausible explanation.
Earth coalesced from primordial dust and gas at 4.5 bya, then the EHB and LHB bombarded it until 3.8 bya, when recently cooled lava formed the first rocks. No Darwinian “warm ponds” of primordial soup existed in those early eras, only a seething, steaming, hellish cauldron of heat.
All across that cauldron, between 4 bya and 3.5 bya, with no precursor forms to set the stage, in corrosive conditions damaging to potential biochemical reactions, a wide array of prokaryotic bacteria suddenly appeared! As if by magic!
 |
If that isn’t miraculous enough, we can take it up a notch. It seems logical to assume that the first life forms had to be of one kind . . . right? It makes sense. First life on Earth, struggling to establish a foothold, it has to be only one kind.
If that were true, “experts” in this field could sleep easily at night. Unfortunately, they can’t because since the late 1970s they have known that not the expected one but two distinct, very different types of prokaryotic bacteria came to Earth metaphorically walking hand-in-hand.
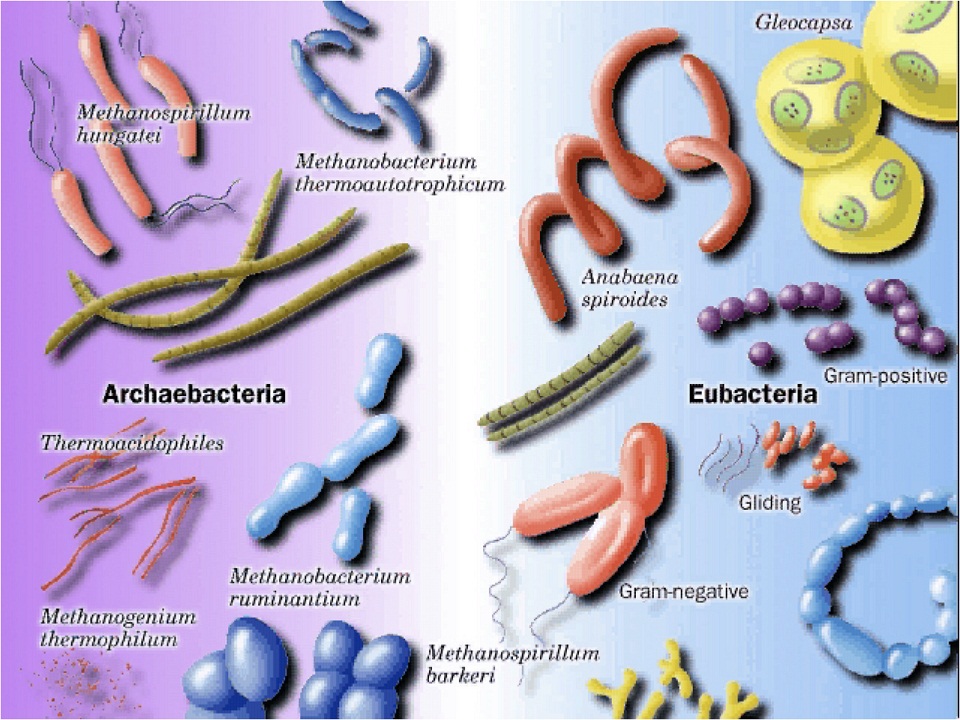
The two types are the archaea (ar-kay-ah) and the eubacteria (true bacteria). As you can see, they are similar, but still markedly different.
The archaea seem older because their functions seem more primitive, so biologists suggest they must have come first and somehow the bacteria evolved from them. However, both first appear in the fossil record at the same time, so arguing for evolution in this case is absurd. It is based on a need for it to be true rather than on facts.
“They couldn’t be that different, could they?” Indeed they could! In 1977, they were divided into the archaea and bacteria because of major differences in the genetics and structure of the two groups. Some parts were similar, but others were vastly different — right out of the chute!
Day One of life on Earth saw two distinct types of prokaryotic bacteria appear at 4.0 bya to 3.5 bya, and leaving the first fossils at 3.5 bya. That means mainstream biology’s official story is a known deception, a fairy tale for willful adults.
 |
No primordial soup in a seething cauldron, no lightning bolts from a cloudless sky. But there was a desperately needed flow of oxygen into the water of the gradually cooling Earth, which would optimize a “terraforming” strategy if that was indeed performed by outside intervention.
Whether or not outside intervention was a part of the equation, Goldilocks Zone protoplanets have certain pressing needs. They require life forms that can endure harsh environments and create enormous biomass (Earth’s bacterial biomass dwarfs all other life combined), live without oxygen, and they must produce oxygen as a byproduct of their anaerobic metabolism.
Almost as if Intragalactic Terraformers were in fact observing our nascent solar system, exactly what the proto-Earth needed was miraculously delivered at exactly the right time in exactly the right way. Prokaryotes were perfect for the job!
Prokaryotic bacteria were the only life forms on Earth for about 2 billion years — 4 bya to 2 bya. They dominated throughout its entire cooling process, until lava rocks mingled with water.
No plants or animals existed yet, not even in their simplest forms. Only masses of bacteria slowly, steadily producing enough oxygen to transform the environment of the protoplanet.
Why was oxygen so important to proto-Earth? Not what seems obvious: “Paving the way for aerobic higher organisms.” That was phase two. Phase one had to be chemically tying up all the free iron available in the newly forming crust.
Iron is abundant on Earth and throughout the universe (most meteorites are dominantly iron). It is extremely reactive with elemental oxygen, quickly forming iron oxide (rust) when the two combine. Thus, until all the exposed elemental iron could be converted to rust, oxygen would not be free for use by any complex life forms.

----------------------------  |
Banded Iron Formations (BIFs), seen above in overlay, are found in every part of the globe and extend back to the earliest crust formations at 3.8 bya. Not until these strata and millions of miles like them formed could proto-Earth host the next phase of complex life development.
Countless prokaryotes generated the oxygen that created the worldwide array of BIFs, but the undisputed O2 champions were the many forms of cyanobacteria, or blue-green algae.
Cyanobacteria seem to be some of the earliest prokaryotes to appear on Earth, usually dated at 3.5 bya, but maybe well before. Whenever they did arrive, they were able to turn water, carbon dioxide, and sunlight into sugar and the oxygen that oxidized the free iron. (They’re still around today in countless forms of blue-green algae).
What is even more “lucky” — what Creationists would call “miraculous” — is that those bacteria and others like them possessed the remarkable attribute of enzymes that prevented their DNA from being ravaged by the hydroxyl radicals that develop during the production of oxygen.
The problem for mainstreamers is that those two diametrically opposed abilities — bacteria creating a poison they were uniquely resistant to — had no cause to develop in a natural way.
From an evolutionary standpoint, how could anaerobic bacteria “gradually” overcome the hydroxyl radicals that develop during oxygen creation, which seems biologically impossible?
Mainstreamers suggest that ultraviolet light striking ice on a primordial Earth could have created peroxide, which could have let certain anaerobic bacteria “evolve” a resistance to it.
This idea, while technically plausible, ignores the fact that ice didn’t begin to develop until a billion or more years after the cyanobacteria.
 |
The ice came with what is known as the Great Oxygenation Event (GOE), at 2.4 bya. It took prokaryotes 1.4 billion years — from arriving at 3.8 bya until 2.4 bya — to make enough oxygen to reduce all of the free iron to the rust in BIFs
The GOE was a catastrophe of immense scale. As soon as oxygen could bubble out of the seas into the atmosphere (see the blue range above), it reacted with methane to create the Huronian Glaciation, the first — and maybe the worst — of what are called Snowball Earth disasters. This is when all, or nearly all, of the Earth’s surface was covered in massively thick sheets of ice.
The Huronian Glaciation lasted from 2.4 bya to 2.1 bya, and no one is sure exactly why it ended when it did. But, by 2.0 bya the planet had been freed of its ice shell, and life was ready to move to the next phase of its journey to complexity.
As if on cue, with oxygen present after the end of the first Snowball Earth, a new form of life appeared to live alongside the prokaryotes — the eukaryotes. To open minds, this could suggest that the Intragalactic Terraformers were also Life-Managers, or Overseers, or Manipulators.
*****
I use the term “Terraformers” as others use “aliens,” which produces negative connotations in most media. “Terraformers” is more abstract and benign. Also, more than one kind can exist.
If they are real, and if they did in fact develop life and/or humans on Earth, then who are they? And who created them? This leads to an endless hall of mirrors echoing the same impossible-to-answer question: Who are their creators???
This brings up what is known as First Cause, the starting place for all of everything — of life and existence itself. No human actually knows anything about it, and we probably never will.
I don’t know, religion doesn’t, and scientists certainly don’t. Yet those institutions are forced by their intense rivalry for hearts and minds to pretend that they actually do know the answer.
Don’t believe either side. Religion and science stand eyeball-to-eyeball and are too frightened to blink, much less acknowledge any doubts or weaknesses in their propaganda. But, luckily, I don’t have those restrictions. I can busy myself trying to discover what is actually knowable.
 |
As with the prokaryotes, many kinds of the new eukaryotes arrived suddenly, and exactly when their much larger, vastly more complex single-cell bodies were capable of thriving in the new environment created by prokaryote metabolism.
Biologists insist this is how classic Darwinian evolution works: when an environment presents an open niche, Nature will fill it. Unfortunately for them, evolution also requires “precursors,” forms of life that provide a base upon which to branch off a new form to fill an empty niche.
Prokaryotes are many things, but they can’t be precursors for eukaryotes. A staggering number of physical and biological differences between them make a direct Darwinian “descent with modification” impossible. It couldn’t happen, especially not “overnight,” which is seemingly how fast they arrived. Yet another miracle!
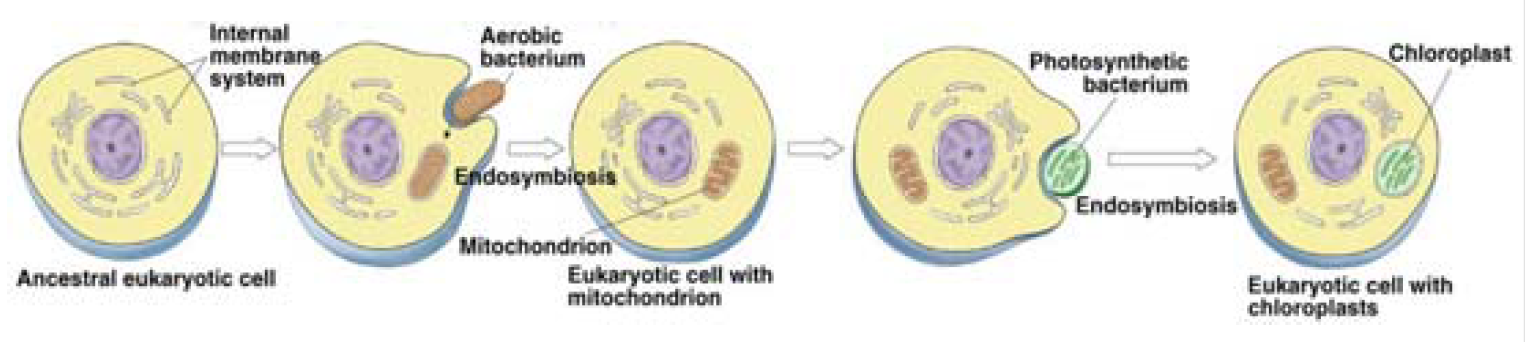
Because direct descent can’t be used, our ever-imaginative “experts” came up with a flight of fancy equal to anything the cosmologists ever concocted to “explain” gravity’s weaknesses.
They suggest eukaryotes must have originated when larger prokaryotes became cannibalistic and consumed smaller ones, turning those into various functioning parts within their bodies, including the crucial, essential mitochondria.
 |
Is that theory likely? Not at all, not even in the ballpark. Understand that a long list of complex differences exists between the prokaryotes and eukaryotes. Size is the most obvious one, but an amazing array of metabolic advances (including an encapsulated nucleus) appeared with them.
Some prokaryotes were larger than others, but their average size would be the light green dots seen in the eukaryote in the illustration at right.
Given so many significant differences, how could larger prokaryotes cannibalize smaller ones, and by doing so grow enormously while utterly transforming their metabolic processes?
 |
A further absurd aspect of the mainstream’s explanation is that all prokaryotes — large and small — managed to live peacefully for 2 billion years, until, out of the blue, some larger ones supposedly turned into cannibals. How? Why?
Even if prokaryotic cannibalism was a known phenomenon (and to my knowledge it is not), wouldn’t the victims be turned into food? Into energy? How could a consumed prokaryote turn into a functioning body part inside what ate it?
This idea is wild speculation, but many strident people with a “Ph.D.” after their names insist it had to occur, and they are not shy about calling on “magic” and “miracles” to make their story plausible. It’s not quite the same as “God did it all!” but it does seem to come dubiously close.
However the eukaryotes appeared, though, they were a huge leap forward from the prokaryotes.
Indestructible anaerobic bacteria (prokaryotes) appeared on the seething proto-Earth as soon as conceivably possible to start transforming the biosphere into something more habitable.
Similarly, eukaryotes appeared precisely at the end of 300 million years of Huronian Snowball Earth, during which the planet’s entire surface was blanketed by ice to a mile or more thick!
Thus, the appearance of the eukaryotes seems timed with astounding good fortune — literally another miracle! — since they arrived precisely when they were capable of thriving on Earth.
| |
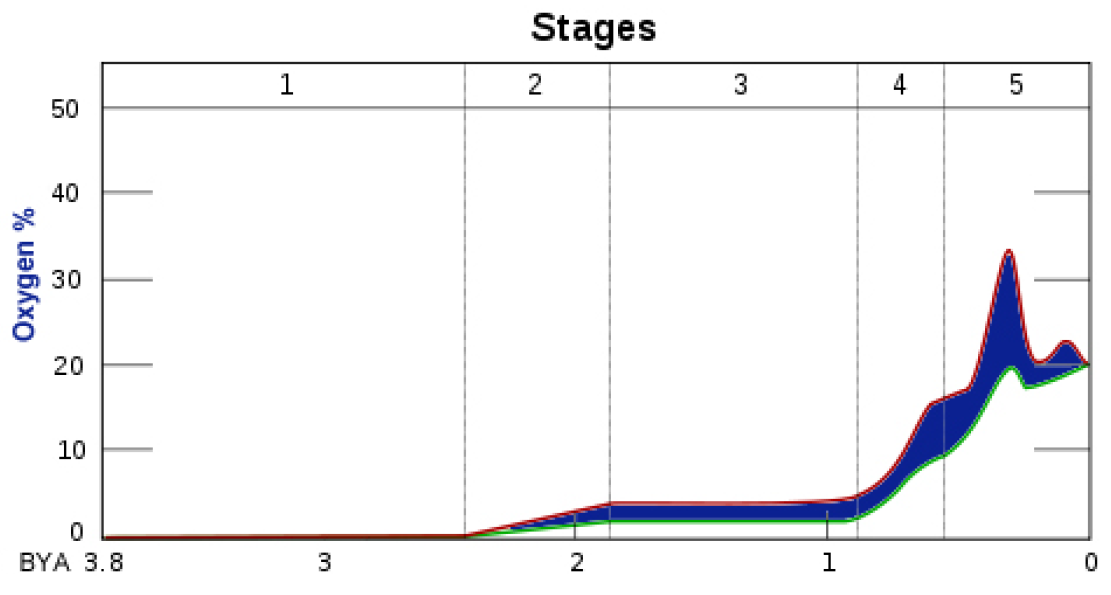
Let’s re-examine the chart at right showing the amount of oxygen on Earth since prokaryotes arrived to begin creating it. Stage 1 shows the 1.4 billion years prokaryotes needed to oxidize the free iron while the Earth’s surface cooled.
Stage 2 had two parts. The first started at 2.4 bya, after the free iron was oxidized into rust and the GOE pumped oxygen into the air to react with methane to create the Huronian Snowball Earth. That ended at 2.1 bya.
The second half of Stage 2 saw arrival of the huge new eukaryotes, which started producing more oxygen than ever before. It ended at 1.85 bya, when prokaryotes and eukaryotes seemed to stabilize in their environments and thrived.
Stage 3 saw oxygen created at a steady rate to do another job as crucial as oxidizing free iron. After the 1.4 billion years needed to do that in Stage 1, another billion years — 1.8 to .8 bya — saw oxygen from prokaryotes and eukaryotes build an Ozone Layer to shield the land and near surface of the sea below the atmosphere.
Complex life cannot exist without that shield. It absorbs nearly all of the Sun’s ultraviolet light, which is deadly harmful to most living species. Unless free iron is oxidized and an ozone layer is established, no protoplanet can ever support complex higher life forms. Another miracle!
Stage 4 extends from .8 bya to .6 bya, and that is the Cryogenian Period, a series of what is now considered to be three separate Snowball Earth episodes, but which may have been one extended 200 million-year-long episode, like the 300 million-year-long Huronian Glaciation.
In Stage 4, oxygen built up under the insulation created by the ozone layer, and with increased oxygen came the next huge step forward in the history of life. Like the bacteria, these creatures appeared at the ideal time, precisely when the environment could support advanced bodies.
 |
|
 |
Stage 5 began at 600 mya and continues to this day. The large upward spike in the amount of oxygen was during the Age of Dinosaurs, when enormous beasts prowled the earth and equally enormous trees and plants covered the land.
This, however, leapfrogs our story.
Let’s return to the next forms of life to join the prokaryotes and eukaryotes. Around 600 mya they suddenly appeared in the planet’s waters and flourished. They were the Earth’s first complex, multicelled organisms — the Ediacaran Biota.
 |
Exactly like prokaryotes and eukaryotes before them, the ediacarans appeared “overnight” in the fossil record in a wide range of varieties.
Just as the eukaryotes followed a global freeze, ediacarans appeared at the end of another — the Cryogenian period, which was the 200 million years (800 mya to 600 mya) that saw either one extended Snowball Earth, or a series of three.
Also as with both prokaryotes and eukaryotes, the ediacarans appeared at exactly the optimum time for them to do so! Yet another “miracle”!
Even more miraculous is that to this day no scientists can say with certainty exactly what ediacarans were or how they lived. They were morphologically distinct from all the prior and later forms, making them utterly unique in the history of life on Earth — a lingering mystery.
 |
Plants … or animals? Nobody knows. They had a variety of shapes — discs, tubes, fronds, bags, even quilts — but did they have mouths? Don’t know. Any digestion? Don’t know. Were they mobile or stationary? Don’t know. There are imaginative interpretations of how they might have looked, but all of those are guesswork.
The key to Ediacaran biota is that in some still unknown manner they seem to pave the way for much more complex animals with skeletons — both internal and external—in the phase of life that followed their enigmatic 50 million year reign as the highest forms of life on Earth.
*****
If we readily challenge conventions, we might accept Intragalactic Terraformers made all this happen as it did — well coordinated and on time. Yet, time is big a problem. How could they take such an incredibly long view of this project?
How could they initiate a process that requires billions of years to complete? The answer has to be “experience,” which means they have no concept of time as we humans understand it.
If indeed they were out and about, seeding life throughout the galaxy, they had to know what they were doing, so projects requiring billions of years would have to be par for their course.
Naturally, mainstream science assures us this scenario could not possibly have happened, but is that opinion based on facts or ingrained bias?
For 50 million years, the ediacarans ruled the seas. The prokaryotic and eukaryotic bacteria still thrived, but in reduced roles compared to the ediacarans, which were vastly larger in size — some fossils were like bathroom throw rugs — and had vastly more complex bodies.
Despite knowing little more about them than their various sizes and their greatly increased complexity compared to what preceded them, we do know when and how their reign ended.
Unlike the prokaryotes and eukaryotes, which functioned perfectly in tandem and are with us to this day, Ediacaran biota seem to have served some purpose and then were eliminated. By 550 mya, they went from clogging the seas to gone.
What happened to them? Why and how did so many go extinct? Nobody understands that. The best answer, really, is: “It just … happened.” Again, like magic, but in a destructive mode.
Regardless of how and why the reign of the ediacarans ended, we know what came next. It was without doubt the most bizarre event in the history of life on Earth — from then until now.
 |
It came with no fanfare, no end of a glaciation, no global catastrophe that left its impact in the fossil record. As with the end of the Ediacaran cycle, the next dominant phase of life on Earth, it is said, “Seems to have just … happened.”
It is known as the Cambrian Explosion, so called because it was a literal explosion of life forms that replaced the multi-cellular, multi-formed, inexplicable ediacarans with a broad range of astonishingly sophisticated creatures.
 |
These are the first “advanced” animals, the first bilaterians, which had left and right sides to go along with the tops and bottoms of the radially symmetrical ediacarans. (Very few of the early bilaterians survive today, the most familiar of them probably the uniquely shelled Nautilus).
We’re talking endoskeletons and exoskeletons; mouths, digestive tracts, and anuses; very large to very small species; predators and prey; and reproduction by male/female sexual relations.
It was a bonanza of new species with enormous complexity. In fact, most animal phyla known today appeared during the Cambrian explosion.
 |
This inexplicable eruption of life was a giant leap toward the myriad of forms on Earth now, and once again the whole of it came seemingly overnight, seemingly “out of nowhere” … a miracle so miraculous, we need a special word for it … a word with Biblical heft and scale:
Supercalifragilisticexpealidocious!
Naturally, mainstreamers struggle valiantly to account for this stubbornly inexplicable event in terms that suit the dogma that only “natural” explanations are acceptable for consideration.
They point out that if the Cambrian explosion required a million years to complete, that is not unduly rapid. Okay, let’s do some quick math:
Prokaryotes appeared between 4.0 and 3.5 bya. Let’s say 3.75 bya. The Cambrian began at 550 mya. 3.75 bya minus 550 mya equals 3.2 billion years with single-celled bacteria and a bit with ediacarans, and then boom! Most animal phyla!
Some say the Cambrian explosion occurred in only a few thousand years. Other insist it lasted a million or more. Compared to the 3.2 billion years that came before it, even a million years is .0003%! In relative terms, it’s an eye blink!
Thus, scientists are left with only verbal fast shuffles and befuddling sleights of mind as they explain a phenomenon that in “natural,” purely Darwinian terms is as humbug as humbug gets.
Mainstreamers are not entirely blind to facts, but they lack other options. Like cosmologists, they must defend an ossified dogma that paints them into a corner where the truth doesn’t fit.
 |
To find any degree of truth about the Cambrian explosion, and all that came before it, we must consider facts at anyone’s disposal, but which mainstream scientists avoid discussing openly.
Prokaryotes, eukaryotes, and ediacarans appear on Earth unexpectedly and rapidly, bearing no apparent relation to each other. But the kicker is that all three arrived at the ideal time for each to perform a critical task needed for turning a raw protoplanet into a viable home for complex life.
Because these facts are undeniable, it really is as if Intragalactic Terraformers were at work, which strongly supports Intervention Theory.
With the Cambrian explosion, the Terraformers seem to have deposited a starter kit with infinite biological potential and flexibility. None of the new Cambrian species had predecessors, yet the planet’s seas were filled with an extraordinarily wide array of animals that to our eyes look as if they must have originated on other planets.
The bleating of mainstream science that all of life must have occurred “naturally,” by means of evolution, can be discounted as the absurdity it is now and always has been. There is no way to account for the sudden emergence of all the forms than to bring up the “A” word — alien.
From Day One through the Cambrian explosion no new life forms had a plausible — much less a certifiable — “precursor,” which is required for evolution to be the force that generated them.
With no clear linkage between early life forms, why can we not consider the glaringly obvious possibility that they all were in fact extrasolar?
Because mainstream science enforces its self-proclaimed “authority” to insist that we can’t.
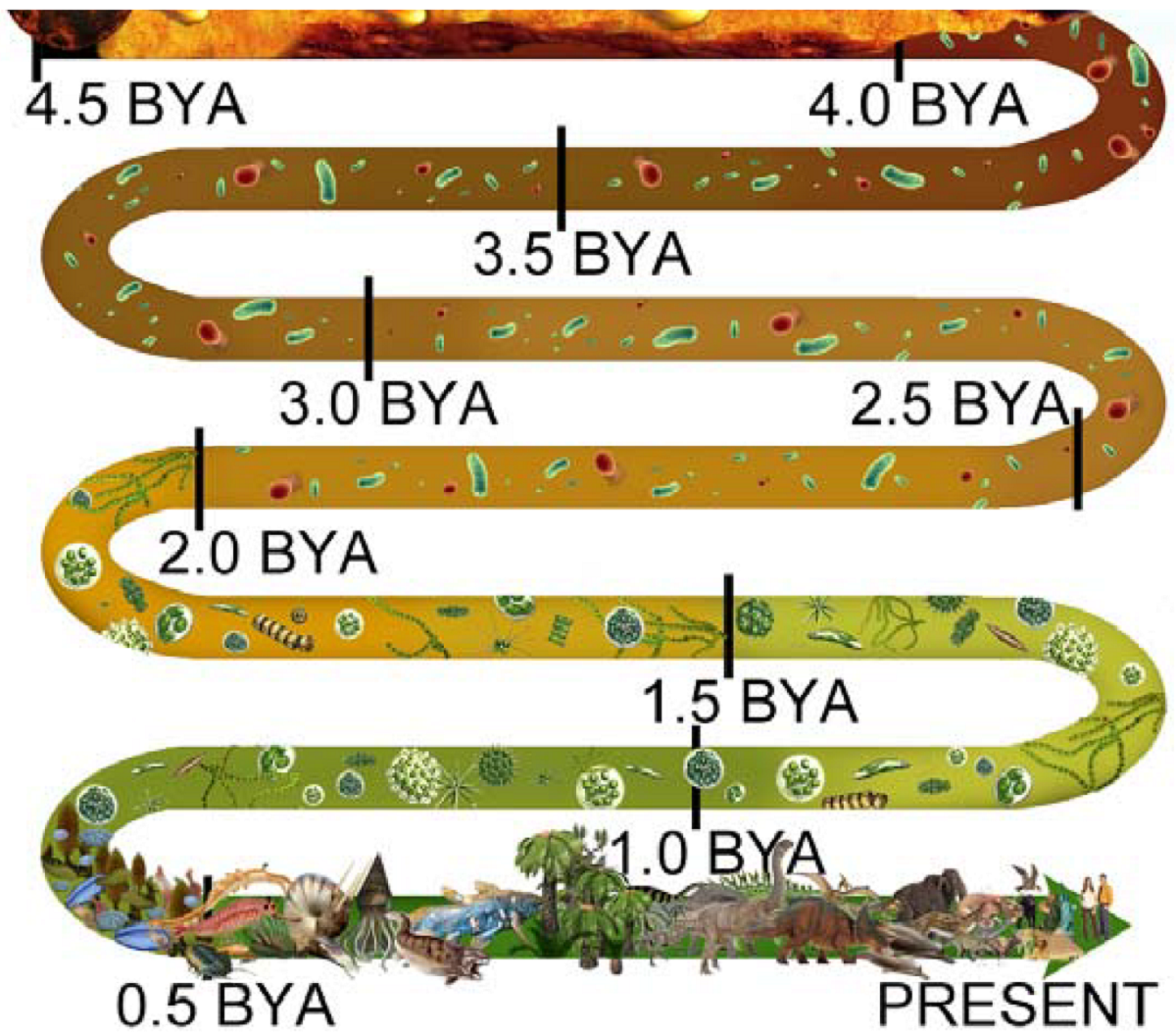 |
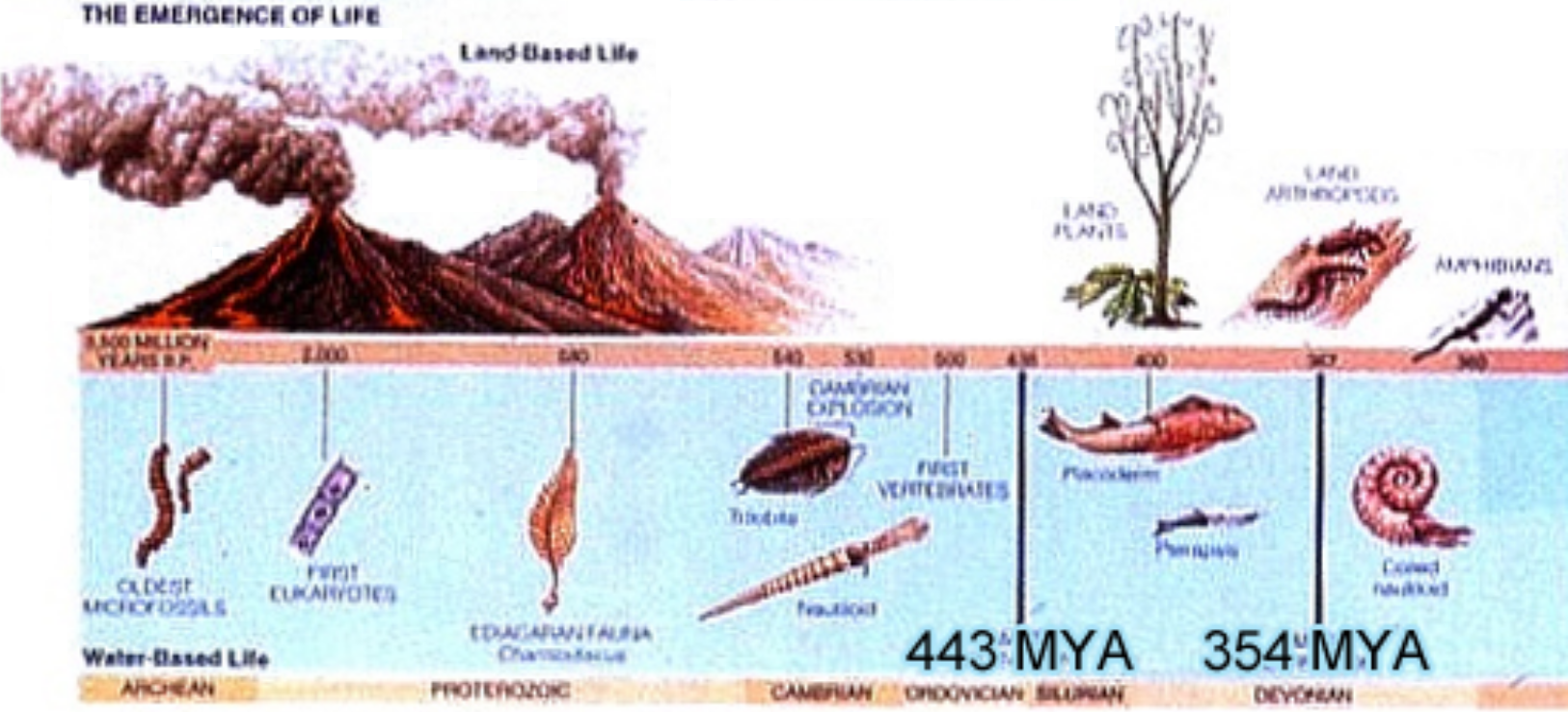
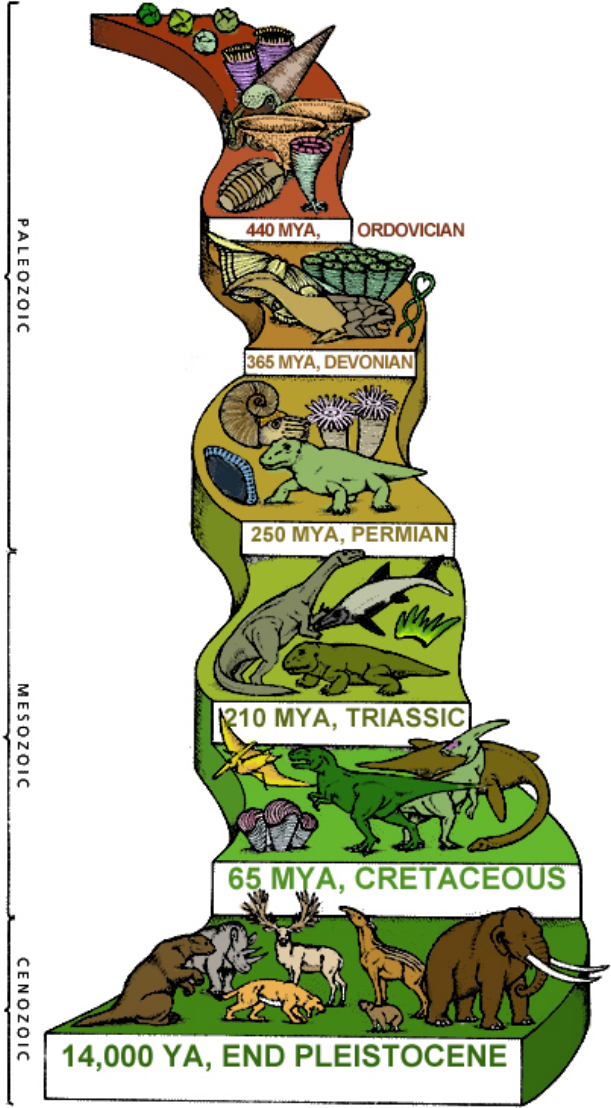
The illustration at right encapsulates the timeline of life on Earth, which we must outline before discussing the far more complex “higher” forms of life that came after the Cambrian explosion.
From coalescing at 4.5 bya, the EHB and LHB brought a hard rain of asteroids onto the Earth until the first crust emerged at 3.8 bya. During this long stretch of turmoil across the seething surface, it was impossible for even the simplest life form to have spontaneously aggregated.
Nonetheless, at 4 bya the first prokaryotes have arrived, appearing “out of nowhere,” hale and hearty, shrugging off all the fire and brimstone pounding down on them as if that hellish siege was no worse than a warm summer drizzle.
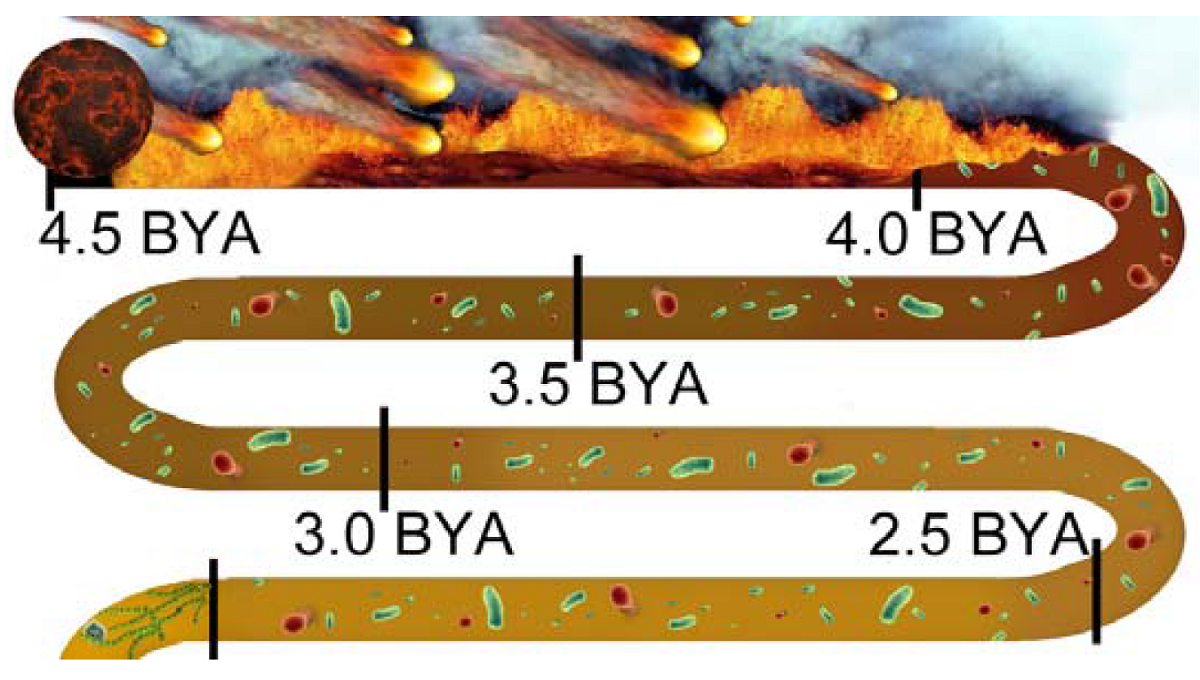 |
Notice, too, that the reddish color representing free iron immediately begins to lighten with the arrival of prokaryotes. This continues unabated with an ever-increasing biomass of prokaryotes through 3.5 bya, 3.0 bya, 2.5 bya, until the next inexplicable “arrival” of eukaryotes at 2.0 bya.
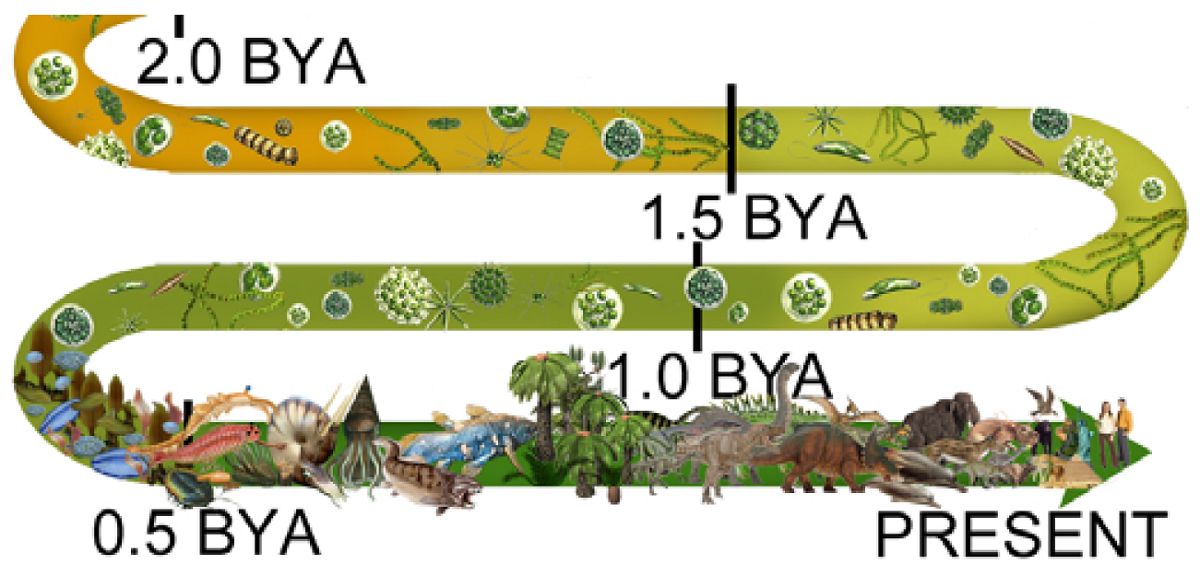 |
From 2.0 bya, the color turns to ever-darkening shades of green, onward through 1.5 bya, to 1.0 bya and on into the last curve of the illustration, where a frond represents the start of 50 million years of world dominance by Ediacaran biota.
Following the ediacarans comes the Cambrian explosion at .55 bya (550 mya), represented by bizarre life forms and something like a nautilus.
Notice how all that remains, the entire panoply of complex life — through dinosaurs and their demise, moving forward to humans — appears and goes forward from only 500 mya, just 15% of the entire 3.3 billion years since prokaryotes arrived. All advanced life is relatively recent.
 |
|
 |
Once again, it seems appropriate to suggest that Intragalactic Terraformers, or entities like them, were making Earth “livable” virtually from Day One, while taking an exceptionally long view of the project from its inception until right now.
Let us recall that the primitive life forms were forged in different crucibles. Prokaryotes came to Earth during an asteroid bombardment. Both eukaryotes and ediacarans followed intervals of profound freeze – the Snowball Earth episodes.
Next came the Cambrian biota, which appeared with no noticeable stress on the planet. It seems as if their time had simply arrived, and when it did, whoever or whatever was in charge of the terraforming decided to bring them on board so they could replace the expendable ediacarans.
Aside from the indestructible prokaryotes and eukaryotes, complex life forms live essentially unchanged until extinction events destroy many species, which are then replaced. Now we will focus on how extinctions shaped our planet.
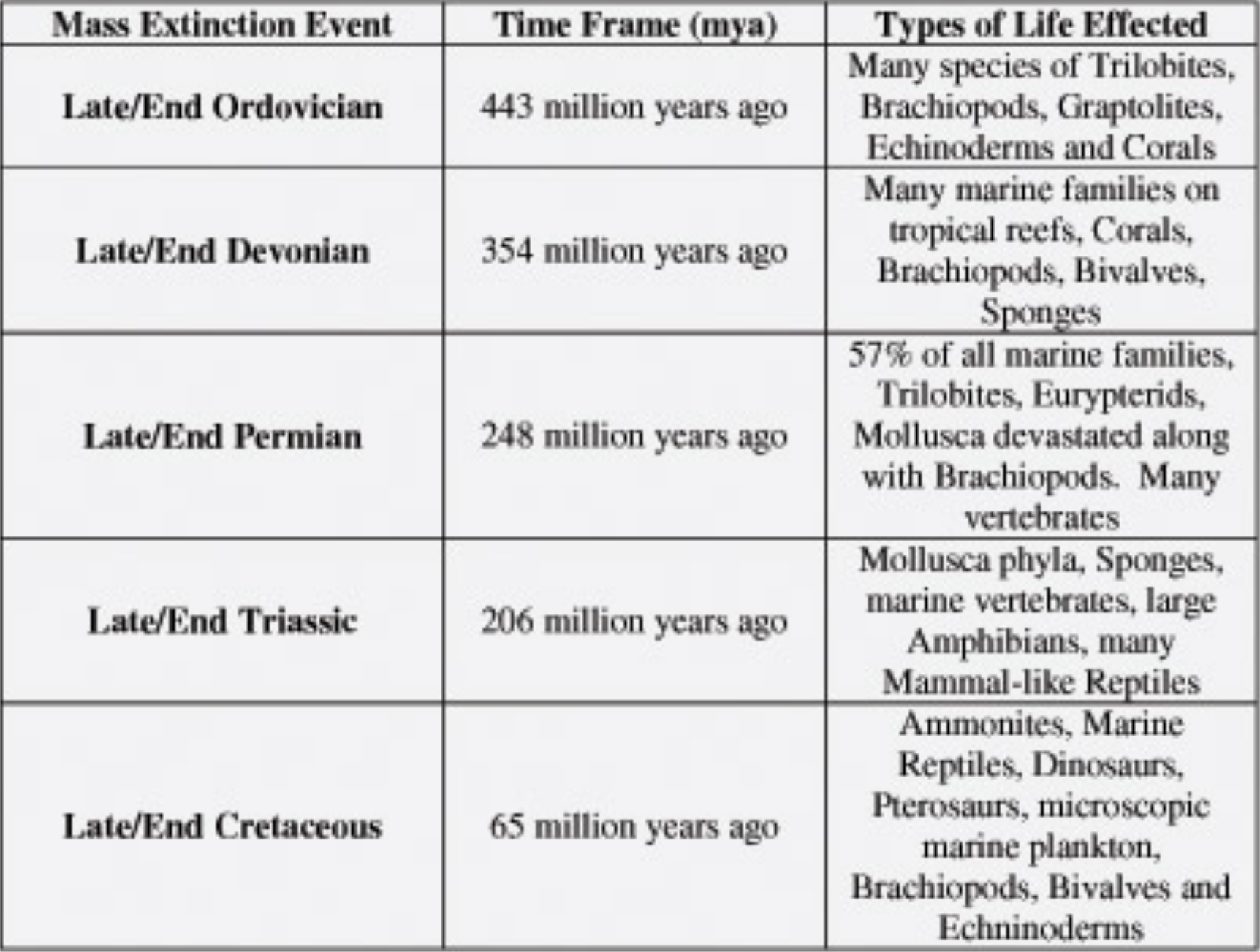 |
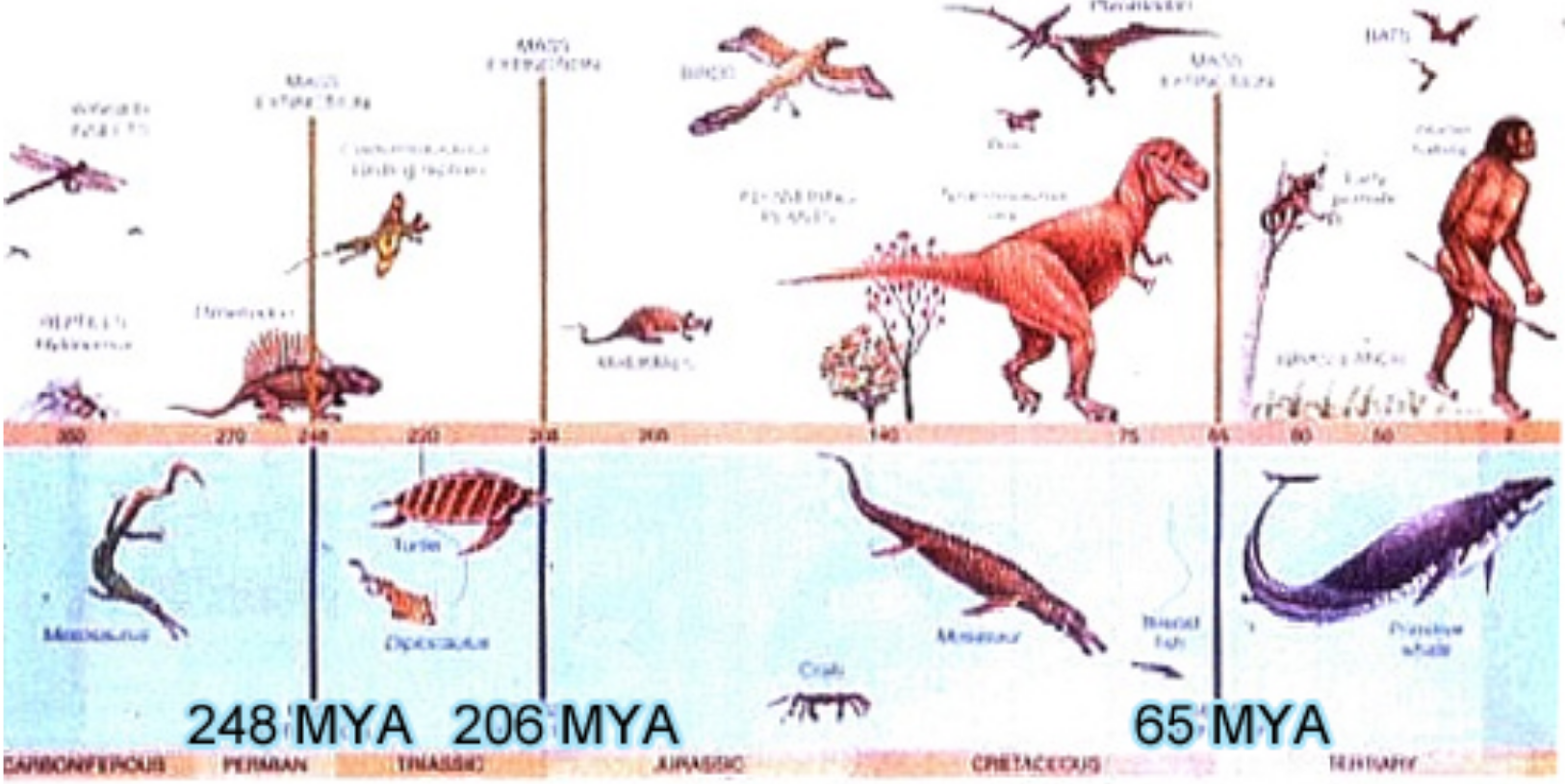
Five major and several minor extinction events have occurred during the 500 million years of advanced life on Earth. As seen above, each of the five is represented by a heavy vertical line.
The chart at right names and discusses salient aspects of the five major events. In each case, the catastrophe had world-wide consequences.
In each case, the era ends with the extinction event bearing its name, then a new era begins.
- Ordovician — 443 million years ago
- Devonian — 354 mya, 89 million years later
- Permian — 248 mya, 106 million years later
- Triassic — 206 mya, 42 million years later
- Cretaceous — 65 mya, 141 million years later
 |
Whether major or minor, each extinction opens up ecological niches that within a few thousand years invariably contain brand new life forms, creatures not seen before in the fossil record.
As always, newbies seem to magically appear, seemingly overnight, which would be accepted as fact if science could afford to consider facts.
During each extinction event, when hundreds or thousands of species are wiped out, others find ways to survive and move into the next era. The environment would be transformed by such an event, sometimes radically, but some of the old species could still survive in it, so they would.
Naturally, the new species that appeared could survive in the changed environment, whatever that might be. Imagine Terraformers analyzing each extinction event, waiting for equilibrium to establish, deciding which new species could survive in the new environment, then making certain exactly such creatures were delivered.
Once again, Intervention Theory provides the most logical explanation for what occurs in the aftermath of each extinction. Equally so, no one can blame scientists for doing what they can to disdain and disparage radical ideas, especially when the ideas make them seem close-minded and even dim-witted. They are not dim-witted.
They simply have an ossified dogma to uphold, and they must defend it without compunction.
 |
Mainstream scientists know that time after time the history of life on Earth shows sudden, “out-of-the-blue” appearances followed by extended periods of stasis, which is “staying the same.”
Stasis is punctuated by extinction events, which some species will survive intact, while others disappear and new species seemingly designed as substitutes materialize to take their places.
Naturally, mainstreamers insist this is proof of evolution where in each case random mutations are “selected” by environmental pressures that result in the modification of a current species.
In the image at right, eight species of Hawaiian honeycreepers show the same kind of genetic radiation that Darwin recognized in finches on the Galapagos Islands. They all remain a twig off the branch of the original species of bird, altering the necessary parts of their bodies to accommodate specific environmental niches.
Eventually, modifications will accumulate in a species until it is altered enough to be no longer recognizable as a member of the initial species that spawned it. It has then become a new sub-species by virtue of those “natural” mutations.
 |
This is microevolution, or “evolution in parts,” a common and well-recorded phenomenon. It was first noticed by Darwin, who extrapolated the idea that it could lead to macroevolution, a gradual change of one species into a distinctly different species, by means similar to micro-evolution, but over much longer time frames.
Macroevolution has never been recorded. No honeycreepers ever turned into finches, or vice-versa. Darwinists have looked for evidence of it since 1859, when Charles Darwin wrote in his landmark Origin of Species that intermediate forms had to be found, or his theory should be abandoned. For 153 years, his followers have diligently sought those ghosts of species past.
Another serious problem with evolution is that not every species will gradually morph into any other form by accumulating positive mutations under the influence of environmental pressures.
The Coelacanth fish ranged worldwide 200 mya (fossil at top). Now it is confined to the Indian Ocean (bottom photo). Thus, it endured tremendous pressures to speciate, but did not.
*****
 |
An interesting double-edged sword wielded by Darwinists is their claim that evolution moves at a grindingly slow pace. As we have noted in considerable detail, this renders evolution null and void as an explanation for the “overnight” appearances of Earth’s earliest life forms. This is an awkward yoke around Darwinist necks.
On the other hand, the glacial pace of change by random mutation is the most critical part of the “proof” Darwinists promote to establish the illusion that macroevolution actually functions. Here is the scenario as they insist it plays out:
Genetic mutations occur randomly among all members of a species. Occasionally, one of those mutations confers some advantage for better chances of survival. Those individuals are “fitter” than their peers, so they have the capacity, and maybe an opportunity, to pass those “better” mutations into their gene pool.
Now comes the “magic” part. It turns out that those macroevolutionary changes in a species’ gene pool occur so incredibly slowly, over tens to hundreds of thousands of years, there is no way to actually confirm them in real time.
“Oops! Sorry! Wish we could oblige you with some solid evidence, but that’s impossible. It happens too slowly. You just have to take our word for it because we’re experts and you can trust what we say.” Or words to that effect .…
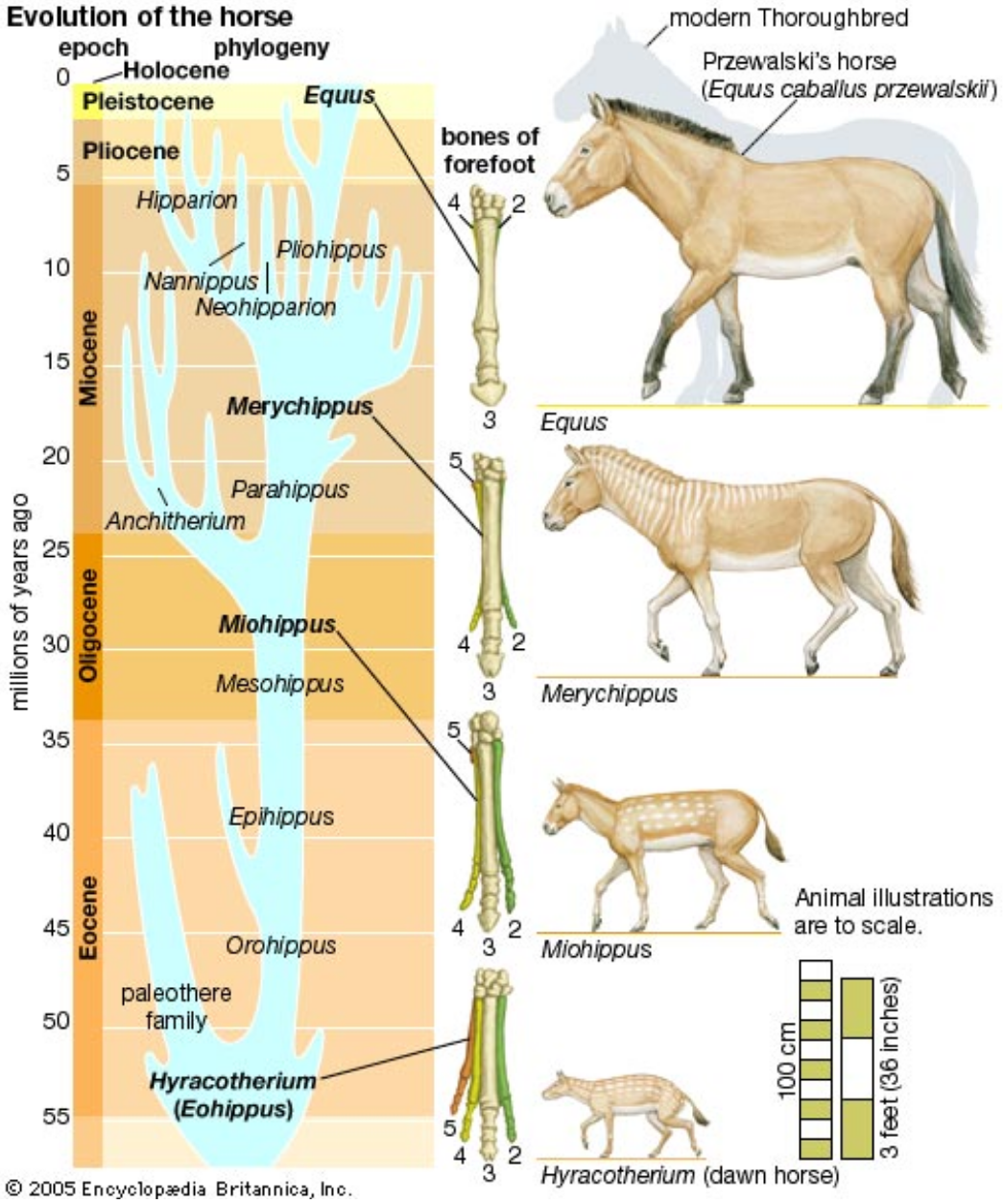
But can we trust them? No, not really. We all have seen a diagram similar to the one above, which the mainstream uses to “prove” horses macroevolved from primitive to modern.
Despite how widely known and accepted these images are, as far back as the 1940s a father of evolutionary science, George G. Simpson, said:
“The uniform (and) continuous transformation of Hyracotherium into Equus, so dear to the hearts of generations of textbook writers, (has) never happened in nature.” And he was right.
 |
In the book Icons of Evolution by Intelligent Designer, Jonathan Wells, he shows how that widely-preached example — and nine more of the most avidly supported mainstream “proofs” of evolution — are complete and utter frauds.
Why is that? Why can’t scientists be trusted to live up to their own hype? To play fair with the facts, and to go wherever those facts lead them? Because they are saddled with a wide range of dogmas passed down from prior generations.
To prop up those dogmas, especially the most vulnerable of them (evolution), science has no choice but to establish and maintain an aura of unquestioned authority. They insist everything had to evolve into everything else, so they bend and twist facts and figures in any way necessary to make that dogma convincing beyond doubt.
I and other Interventionists do doubt, sincerely and profoundly. We take to heart the words of Don Marquis: “When people think you make them think, they will like you; but when you really make them think, they will hate you.”
With the ease of access to information via the internet, there is little excuse any more for not actually thinking, and fact checking, whatever science proclaims as their “received wisdom.”
 |
 |
 |
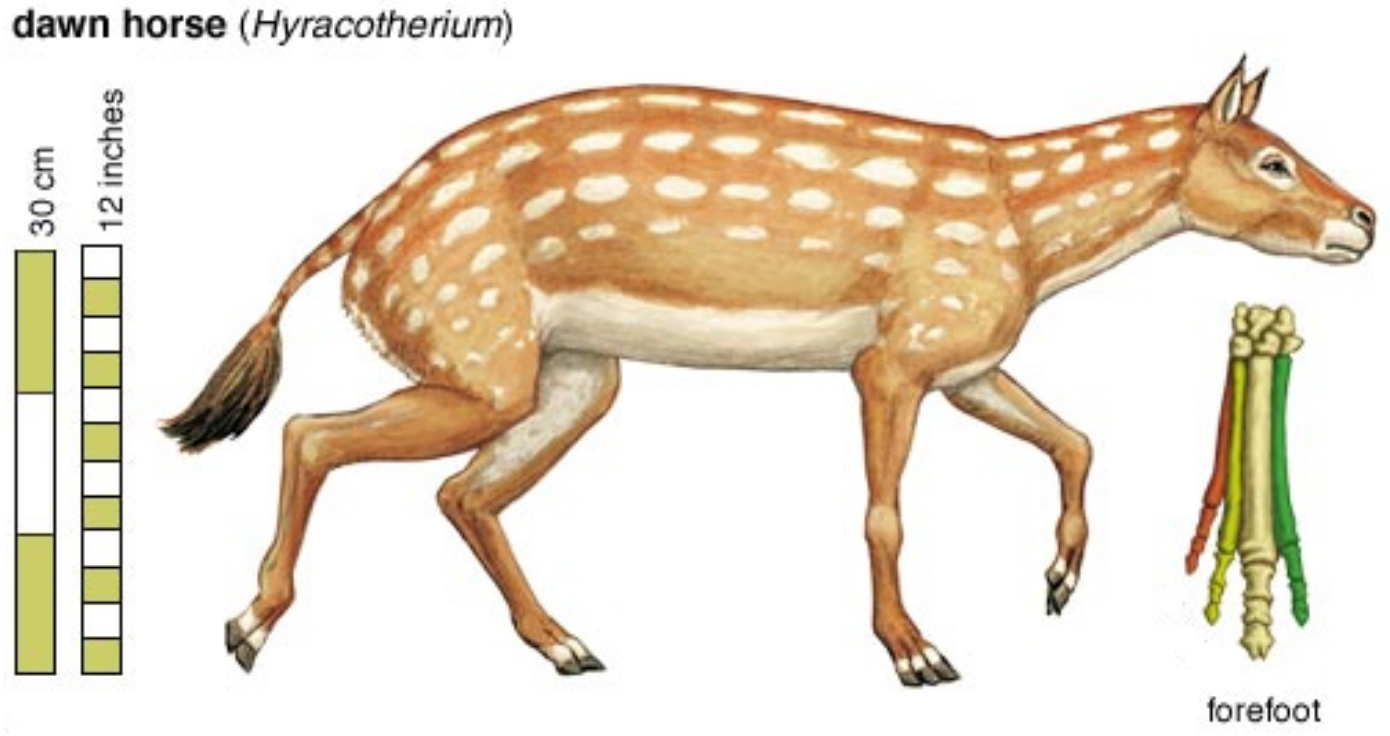
With that chide in mind, let’s take a closer look at the “evolution” of the horse proposed above.
Meet Hyracotherium (Hyra), “dawn horse,” supposedly the root stock of all horses, living from 55 to 35 mya. Below is its full skeleton.
The mainstream concocts hooves that clearly are imaginary, then gives it a horse-like outer covering, and bingo! A proto-horse is created!
Who can say with any authority that Hyra did not have an elephant’s feet and a rhino’s hide? Like beauty, horse is in the eye of the beholder.
In the case of horses, there is no real evidence that any one type of pre-horse was transformed by means of macroevolution into any other.
Mainstreamers never hesitate to bolster their case for evolution with misleading illustrations like the ones for the horse. Those deceptions are designed to make the uninformed think the dogmas are based on solid facts. They are not.
Furthermore, any time a species is altered by genetic manipulation, no matter how radically it is “mutated” (say, legs growing where antenna belong, or vice versa), if it is allowed to “breed back” into its native population, in only a few generations the new mutations are eliminated!
Thousands of similar genetic studies show that every species has a norm — a range of variation. A horse can be from a Shetland to a Clydesdale, but not the size of a Chihuahua or an elephant.
Nature adheres tenaciously to physical and/or biological norms, such that any mutations — positive or negative — tend to be eliminated by the collective “wisdom” of the gene pool.
Despite those problems, mainstreamers insist evolution occurs too slowly to leave visible traces, so it can never be analyzed, confirmed, or denied. That renders it immune to criticism, much less negative proof. You can’t hit what you can’t see, and no one can see this humbug.
Just like cosmology’s “fudge factors” of dark matter and dark energy, Darwinists hide their “missing” evidence for macroevolution behind an equally dark veil of obfuscation — or bluster when anyone dares to challenge their position.
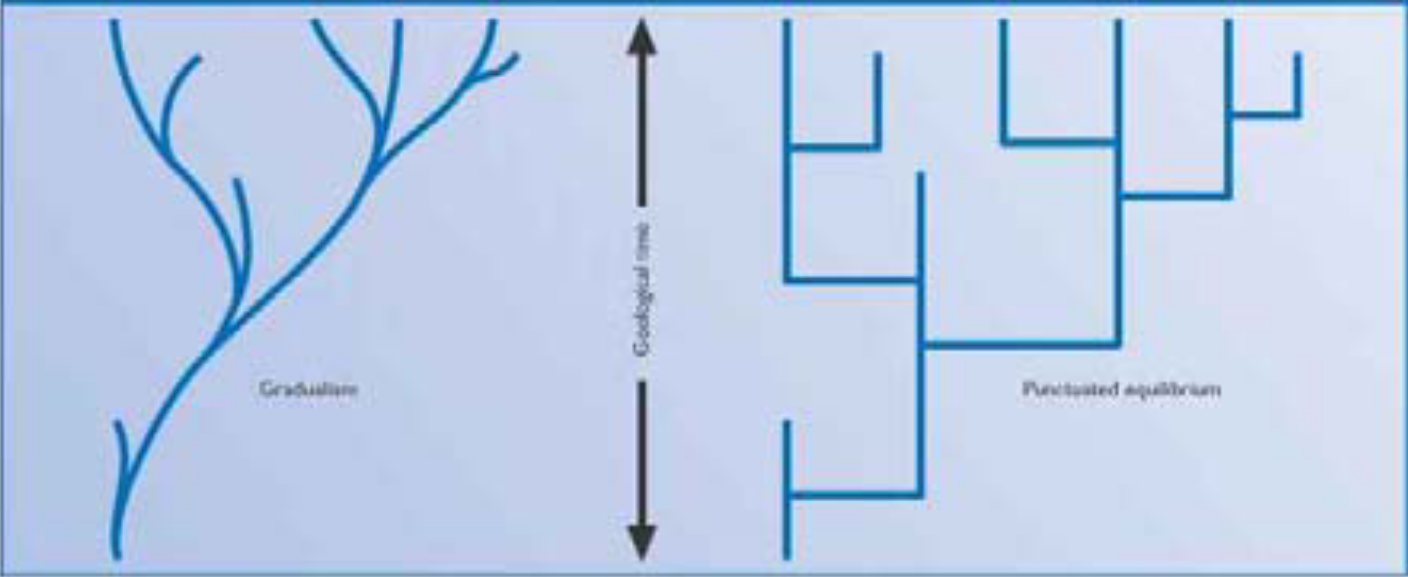 |
Despite the reality that microevolution does occur, Darwinists still confront the intractable problem that since Day One each new form of life has appeared on Earth so rapidly that in relative terms it can be considered overnight.
To plug this hole in evolutionary logic, clever Darwinists concocted a theory of explanation that could be construed to make sense in natural terms. It is known as Punctuated Equilibrium, or as its many critics refer to it, “punk eek.”
This illustrates gradualism (left) and punctuated equilibrium (right). Gradualism says evolution proceeds by the steady accumulation of small genetic changes over extensive periods. Punk eek sees morphological change concentrated in “brief bursts” that produce many new species.
Purist evolutionary theory strongly supports gradualism, while reality supports punk eek. Mainstreamers regularly debate which is true and which is based on misguided ideology.
Misguided or not, punk eek’s ideology is that when evolution can proceed via gradualism, it does precisely that. However, when any crisis puts evolution under intense external pressure, somehow it can press an accelerator and rapidly modify any life form into something markedly different to fill one of many vacuums Nature is alleged to abhor — an empty ecological niche.
Punk eek’s real purpose is to deflect awareness that life comes to Earth suddenly, too fast to be accounted for by gradual genetic modifications. It is the acknowledgement of a glaring fact that its critics want to keep pretending doesn’t exist.
To that end, another clever group of “experts” have concocted a different but equally clever way to explain the sudden appearances of life on Earth, and the equal suddenness of life’s many profound physiological transformations.
In 1818 a French zoologist noted that vertebrate animals seemed to be flipped-over invertebrates showing no transitioning from one to the other. His theory was ignored until recent molecular analysis provided great weight to his argument.
From molecular analysis has come Modularity Theory, which states that all speciation results from wholesale mutations in genetic modules, the general parts of nearly every animal body: head, trunk, fore limbs, hind limbs, digits, etc.
Modularity theorists acknowledge the absurdity of arguing that invertebrates could have become vertebrates by accumulating random mutations. They suggest life’s sudden transformations can best be explained by mutations in entire genetic modules, which is rapid macroevolution rather than extensively sustained microevolution.
Like the primordial soup theory, punk eek and modularity theory are audacious claptrap that is offered because it is better than facing the truth.
*****
 |
Against whom or what are scientists compelled to defend their brazen scenarios? Who provides the pressures that cause them to botch so many attempts in their alleged quest for truth? Those crafty Creationists and Intelligent Designers!
For decades Creationists and IDers have battled for the hearts and minds of those convinced that “science has all the answers,” and that science has especially valid answers when the questions deal with the origins of life and of humans.
Naturally, and obviously, Intervention Theory does not yet figure into the battle because our status as contenders is uniformly disregarded. However, while at present we are few, we take a long view, knowing that facts and reason are solidly on our side, and ultimately we will win.
In the fullness of time, as vehicles of change like this eBook fall into more and more hands and hearts and minds, the other contenders will be forced to release their strangle holds on each other to focus their undivided attention on us.
We welcome this and anticipate it, knowing the end game can only benefit every human on the planet. Why? Because a basic Interventionist tenet is that we cannot responsibly chart our course into the future until we are certain of the true provenance from which we emerged.
Only then can we make choices about our lives and about our futures that are best for all of us, rather than being driven toward emotional dead ends and intellectual cul-de-sacs by polarizing pressure groups like science and religion.
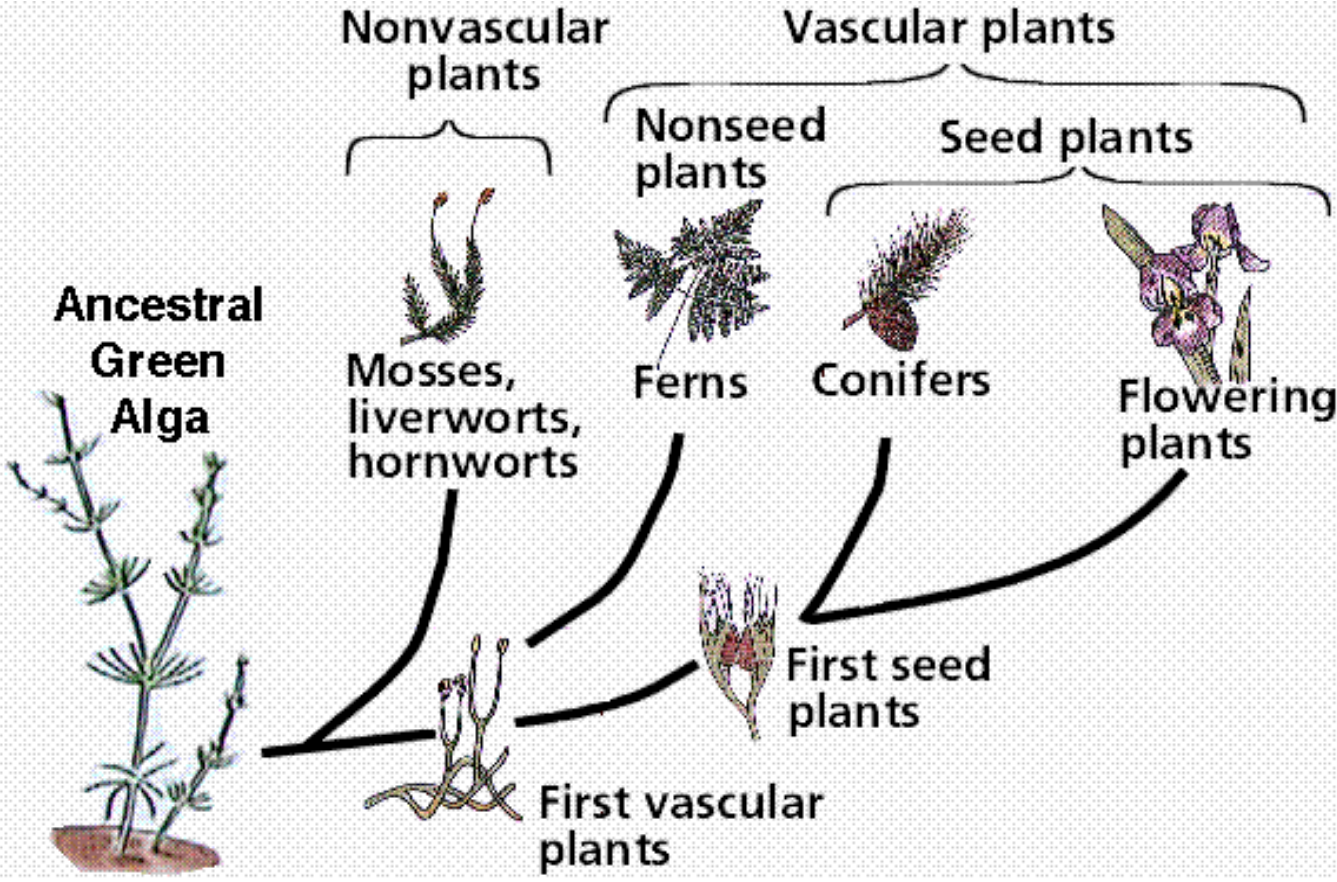 |
Roughly 500,000 plant species exist today, with 300,000 green plants, and 80% to 90% of those flowering. Botanists say they all evolved from one-celled algae. In fact, they claim that of the four types of algae that moved from the seas to inhabit the land, only one of those managed to spawn all 500,000 species of land plants!
How could that one species of algae endure as thick mats on the land from around 1.2 bya to around 450 mya, then seemingly on cue initiate a transformation into the bryophytes, lycopods, ferns, and then the very complex gymnosperms and angiosperms we are most familiar with?
This is what we’re asked to believe — that four forms of algal mats blanketed the land from 1.2 bya to 450 mya (750 million years!), then one suddenly gained super powers and abilities far beyond those of mortal algae, spawning myriad new forms while its three amigos stayed mats.
This was another of those magical moments in history where something … just happened! It was another of the “God-worthy” miracles that always seem to occur on Earth at exactly the right moment in exactly the necessary way.
The arrival of the flowering plants remains an intractable difficulty for mainstreamers to try to explain. Even Darwin was flummoxed, calling the flowering plants “an abominable mystery.”
Although the fossil record since Darwin’s time is greatly enhanced and filled in, there is still no clear path from gymnosperms to angiosperms. Then, along with those flowering angiosperms, came the insects, which are equally puzzling.
 |
Flowering plants appeared on Earth around 130 mya. With them came insects, which today are trillions strong and provide incredible tonnage of biomass. Together they comprise 5/6 of all the animal species alive. How did that happen? How did insects come to dominate the planet?
From 400 mya to 130 mya (270 million years), only a few insects existed, and they spent their lives foraging in detritus on forest floors. That was 270 million years living as foragers, then flowering plants appeared from out of the blue.
When that happened, a genetic “bomb” dropped onto the foragers, making them proliferate like weeds to coexist with many types of flowering plants. It was yet another of the “miracles” that are so implausible, yet so frequent, in Nature.
This could be considered a classic example of punctuated equilibrium, except it lacks enough external pressures to justify its occurrence. No major catastrophes happened around 130 mya.
 |
As with the Cambrian explosion, flowers came when they did because the time for them seems to have been judged right. And who could make that judgment? Only Intragalactic Terraformers.
As the flowering plants multiplied and spread across the planet, insect species increased and spread along with them, forming the symbiotic relationships that define their existences today.
With so much species proliferation occurring relatively recently, it would seem that hundreds and thousands of intermediate forms would be easily detectable in the fossil record or — more convincingly — among/between living species.
Despite that undeniable reality, and to the great chagrin of mainstream scientists, no discernable evidence of “natural” evolution can be found among/between flowering plants and/or insects. [The famous Peppered Moth case of observable “evolution” is one of the 10 mainstream cases proven fraudulent in Icons of Evolution.]
With what would seem to be literally millions of opportunities to isolate some transitional forms — say, a housefly becoming a firefly — scientists remain shamefully empty handed.
This is yet another convincing indication of Intragalactic Terraformers passing through the early Earth’s neighborhood again and again, coordinating the development of life here — from beginning to end, every species alive.
I understand how radical that sounds, and how unlikely it seems, but it does fit the facts better than any theory put forth by the mainstream.
 |
The last major extinction ended the Cretaceous period at 65 mya. That one is familiar to nearly everyone because it wiped out the dinosaurs.
Our purpose is not to dissect this event, or any life patterns between the start of the Cambrian explosion and the end of the dinosaurs. This is available in books, articles, and periodicals that anyone with curiosity about it can easily obtain.
Our point is that Intervention Theory suggests Stock Species (those appearing on Earth with no predecessors) always seem to arrive when the timing for it is most opportune, when they can survive in the environment of the moment.
During the long stasis intervals that follow the extinction events, stock species can — but don’t necessarily (as with coelacanths) — microevolve to radiate new related species (honeycreepers).
Understand that environmental variations could be drastic. In the Triassic period preceding the Jurassic, oxygen is estimated at 80% of levels today. In such an atmosphere, only relatively small animals and plants could survive.
After the Triassic extinction led to the Jurassic and Cretaceous periods, the atmosphere went in reverse, to an oxygen content 150% more than today. This would permit significantly larger plants and animals, and soon they appeared or radiated from whatever survived the extinction.
The earliest dinosaurs were, in fact, part of the Triassic, seeming to begin small in the reduced oxygen content of 230 mya. However, after the Triassic-Jurassic extinction produced that new atmosphere supercharged with vastly more O2, plants and animals grew to gargantuan sizes.
 |
 |
At right is a petrified piece of wood from a forest of enormous trees that once covered the terrain of what today is South Dakota’s “Black Hills.” In the Cretaceous period of 120 to 130 mya, these trees towered 500 to 600 feet, or more, and were as wide and thick as a jumbo jet.
Scurrying among such monstrous trees, trying to avoid being squashed or consumed by any dinosaurs, were small furry creatures that had appeared around 200 mya as the early forms of mammals. Some might have been stock species brought in not long after the prior extinction.
 |
After the Cretaceous event, new stock species might be brought in, too, from which countless larger mammals could radiate in the ecological niches left by the extinction of the dinosaurs.
With such a drastic reduction in oxygen levels (2/3 of the estimated Cretaceous amount), many new species had to appear which were adapted to that amount, and, as always, they did appear.
Intervention Theory suggests that the new stock species were starting points for microevolution to do its work, as in the cases of the Hawaiian honeycreepers and Galapagos finches (below).
 |
How do the stock species arrive? Obviously, they were delivered. By whom? Intragalactic Terraformers are a solidly viable option, but Creationists and IDers will insist all of it is the handiwork of God, while any Darwinist would howl in protest and insist that the idea of stock species being delivered to Earth is blasphemous to the “sacred” dogmas they profess to believe.
Who is right? Who is wrong? That will surely be established in the fullness of time. For now, each individual can only decide for themselves whom to trust and what they prefer to believe.
 |
The Cretaceous extinction was followed by the Cenozoic era, which extends from 65 mya until now. The Cenozoic contains three subunits, one of which is the Miocene epoch — 23 to 5 mya.
For humans, life doesn’t get interesting until the Miocene, so we must examine how it unfolded. The Miocene is so important because it is when Miocene apes became a dominant species.
The mainstream experts responsible for the line charts at right — paleontologists and geneticists — work with an astonishingly small amount of evidence to make the kind of bold and specific proclamations they assert. Of course, each bold claim can change (and often does) with the next bone shard or tooth chip found in different eras, but for now, this is how they see and preach it.
The first steps on the ladder to humans are the “proto”-primates that radiated widely after the Cretaceous extinction event. Interventionists say some of these were stock species, although no “expert” would give that idea any credence.
 |
Next came the earliest prosimians, which were considered to be primates but not yet classified as monkeys, although they are monkey-like in many ways. Several existed in the Eocene, but only three have survived to modern times. They are the galago/bushbabies, tarsiers, and lemurs.
 |
 |
The next early primates, monkeys, appear at around 33 mya. They control their niches for the next 10 million years, until the start of the Miocene, when the first tailless apes appear.
With each of these creatures, Darwinists argue, one led to another like climbing a ladder. In the proto-primates, they declare, genetic mutations slowly accumulated to such a degree that in the course of 10 million years (65 mya to 55 mya) one or more species evolved into prosimians.
Superficially, this sound quite plausible. After all, 10 million years is one heck of a long time; enough, it would seem, for virtually anything to happen, especially since we see microevolution occurring in isolated breeding populations that adapted to local conditions by radiating from stock species in only several thousand years.
On the other hand, coelacanths make clear that microevolution doesn’t necessarily occur at all, even if environmental pressures are intense. So it is possible that some stock species come here from places where microevolution can’t occur, while stock species from other places will have ranges within which they readily microevolve.
No matter whose beliefs about this are correct, prosimians either transformed into monkeys by gradual changes, or stock species of monkeys were delivered and others radiated from them.
 |
However it occurred, after 22 million years as Earth’s dominant primates (55 mya to 33 mya), prosimians gave way to monkeys and rapidly faded into obscurity, thriving today only in the very few places where monkeys do not inhabit.
Next, by either of the two processes described above, transformation or terraforming, after 10 million years of monkey dominance (33 mya to 23 mya), the early Miocene apes appeared and took over primate ecological niches as deftly as the monkeys took them from the prosimians.
With the appearance of the Miocene apes, the squabbling over human origins gets cranked up. Which Miocene ape, or apes, led on to gibbons, gorillas, chimps, and, finally, to humans?
 |
The Miocene is alleged to have been crawling with apes (no pun intended). Scientists claim as many as 100 different species lived in the Old World, but none ever lived in the New World. Monkeys, though, are native in South America.
How could so many apes flourish so widely, then find themselves whittled down to the four main groups we have today: 3 types of chimps, 2 of gorillas, 2 of orangutans, 4 of gibbons?
Is it possible someone has made a mistake at some level? Might there have been only, say, 30 to 50 Miocene species rather than 100? The answer is yes, definitely, and here is why:
In the field of anthropology, as in most other fields, fame equals status, power, and money. Therefore, careers in anthropology depend on making big discoveries or naming new species.
This explains the wide range of species names in the array of humans, pre-humans, could-be-humans, or maybe-someday-will-be humans.
 |
Entire species can be, and have been, created from as little as variations in size and position of cusps in fossilized teeth. And that can be in only one, or a few, teeth isolated and alone, not multiple teeth attached to a jawbone or maxilla.
Every anthropologist struggles mightily to find some fossilized tooth or shard of bone that can, in any clever or roundabout way, be interpreted as a “new” species. If they manage to pull that trick off, their careers can blossom overnight.
The point to keep in mind is that while clearly there were a large number of Miocene apes, the odds are quite long against the number actually being the 100 professed by so many “experts.”
*****
 |
Whether any Miocene apes survived to become the apes we have today, or went extinct at some point, modern anthropologists insist that when on the ground they moved as quadrupeds, and when in trees they brachiated (swung by the arms) or meandered their way atop limbs.
Today, the dominantly brachiating apes are the gibbons and the orangutans, whose arms are so long relative to their bodies that when on the ground they tend to hold them up and walk bipedally. All chimps and gorillas can walk bipedally, and often do, but their preferred method of ground travel is quadrupedally.
Given those facts, would anthropologists allow for the possibility that any Miocene apes were consistently bipedal? Could any have walked on two legs in a manner similar to humans?
NO! Not in a million years! Better yet, not in the 23 million years since the Miocene began!
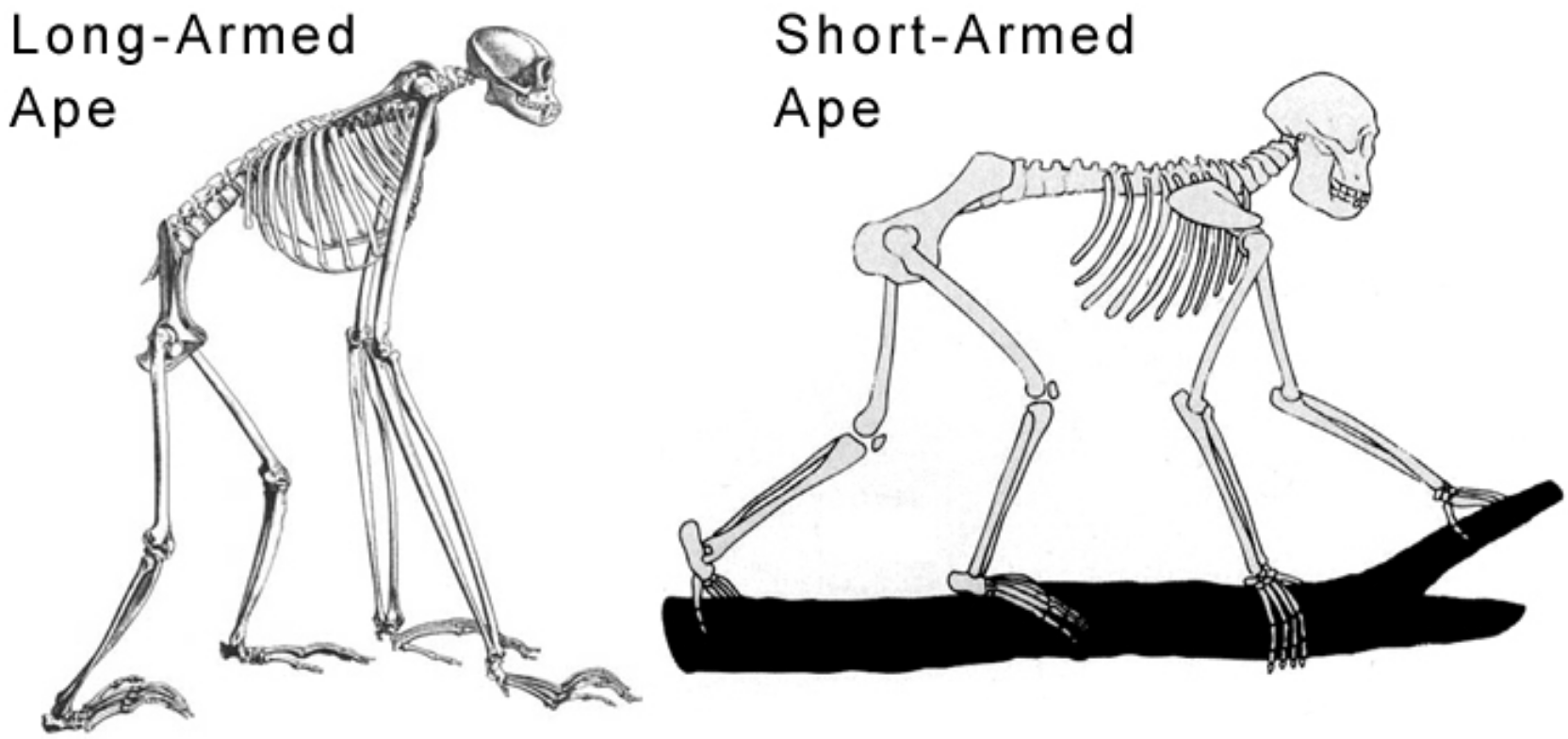 |
Darwinist dogma asserts that apes could only evolve into humans by gradually transforming their genetic code during millions of years by a fortuitous accumulation of positive mutations.
This is despite the two most significant changes that distinguish humans from apes — bipedality and increased brainpower — appearing in every respect fully developed in the fossil record.
Bipedality is especially puzzling. If apes did not gradually rise up off all fours to become bipedal humans, where might the many ancient bipedal creatures called pre-humans have come from? The Miocene epoch is the place to start looking.
Among more than 20 genera of apes, and 50 to 100 species, were examples the size of midgets, humans, and giants. However, only four genera survive today — those four quadrupedal apes — gorillas, chimps, orangutans, and gibbons. Why and how did so many go extinct? Or did they?
Two basic types of Miocene apes existed: long-armed and short-armed. The long arms were longer than their legs. Modern apes are of this type because quadrupeds need arms longer than their legs to move on the ground in comfort.
Short-armed apes are a mystery. Their arms are about the same length as their legs, so when on the ground, their hands would dangle near their knees. That sets them apart from humans, since our legs are considerably longer than our arms.
 |
 |
The short-armed apes could never have moved comfortably down on all fours, nor would they be good {tip content="Brachiators use their arms to move from tree branch to tree branch, through a process called brachiation.Their arms are longer than their legs, and are much more powerful."}brachiators{/tip}. This has generated serious debate about how well they might have moved their bodies when up in trees or on the ground.
If they were fully arboreal (living in trees), their short arms would be unlikely to brachiate their bodies easily or well. They would be far more likely to move as in the photo at right, much like monkeys that move along the tops of branches.
The problem for apes is that they lack crucial elements required for safe branch scampering:
- a highly flexible tail to act as a balancing mechanism;
- shoulder blades on the sides of their torsos rather than along their upper backs; and
- apes are much heavier than monkeys.
Balancing a 20 to 100 pound body — or more! — on branches of any size would be quite a trick.
At right is a chimp that lost its hair due to a skin disease, but it shows how densely muscled they are under their hair. In fact, pound-for-pound they are 5 to 10 times stronger than any human. Thus, to say short-armed Miocene apes moved by shuffling along branches seems farfetched.
 |
 |
If short-armed apes were too large to scamper along the tops of limbs, and their arm length and shoulder design made brachiation at most an occasional mode of travel, is it possible they might have preferred living on the ground?
Yes, it is quite possible. In fact, all apes spend at least a part of their time on the ground. The extra-long arms of the brachiators make ground travel a small part of their movement repertoire, but quadruped gorillas and chimps comfortably split their time between trees and the ground.
Mainstreamers have decided that when short-armed apes were on the ground, they must have moved as seen at right, shambling in a bent-knee shuffle below a stooped upper body, swinging their knuckles near the ground. This is where the term “knuckle-dragging” comes from.
This oafish, shambling movement is much like Groucho Marx’s famous caricature of human walking. However, notice how little effort they would need to stand a bit straighter, then a bit more, until soon they would be fully upright.
It is difficult to imagine that during several million years not one of those creatures ever thought to do that. It would allow vision in a much wider panorama, and also relieve a great deal of stress on their low back and buttocks.
Even cartoonists know the mainstream position is absurd, so let’s join the one above assuming at least one — and no doubt more — short-armed, knuckle-dragging Miocene apes made a radical decision to improve their world in a number of very useful ways simply by standing upright.
***
 |
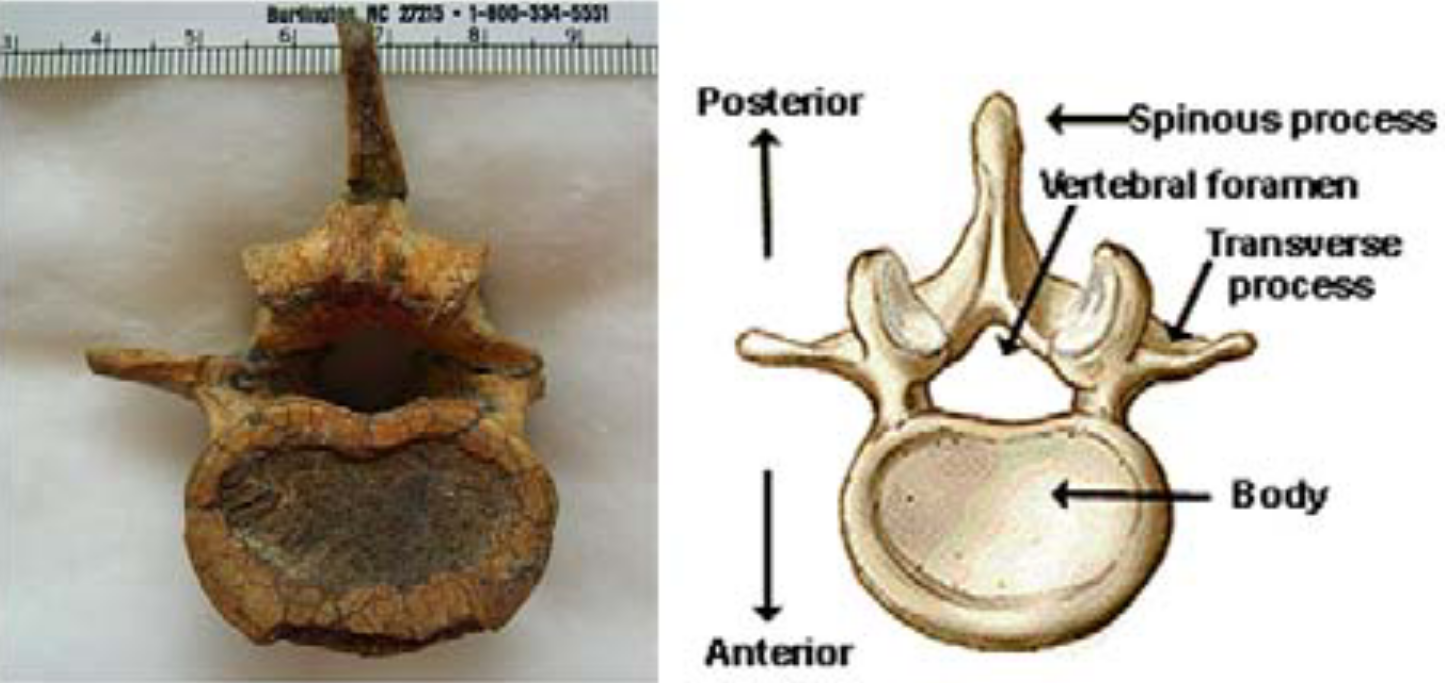 |
A long-established fact in anthropology that its practitioners dutifully ignore is this: bipedality seems to have appeared early in the Miocene.
In 1902, Sir Arthur Keith proposed the idea that the evolutionary ancestors of humans and apes must have been bipedal, and that modern apes developed specializations to move on all fours.
Keith based his theory on observations gained as a surgeon, which dealt with the arrangements of primate and human internal anatomy, rather than the bones and muscles of anthropologists.
Unfortunately for Keith, his data contradicted the “received wisdom” of his peers, which was that ape evolution required a gradual “upward” progression from the quadrupedal apes to fully bipedal humans. By the middle 20th century, no one seriously doubted the mainstream scenario.
Then, in 1961, fossil bones of an early Miocene ape were uncovered on the slopes of the Moroto volcano in eastern Uganda. They dated to 21.6 mya, near the Miocene’s beginning at 23 mya.
The owner of those bones came to be known as Morotopithecus, without doubt one of the first arrivals among the tailless primates that would rapidly take over from the dominant monkeys.
Among Moroto’s fossils was a spinal vertebra (at right) astonishingly like the rigid vertebra of all higher primates, including humans. It was so much like a human vertebra (below right), it seemed designed to support efficient bipedality.
No one knew what to make of this “misplaced” bone that had no business existing at 21.6 mya. As with all pieces of new knowledge that, when revealed, contradict ossified scientific opinions, this one was quietly shelved for two decades.
 |
In 1981, it was noticed by Aaron G. Filler, an anthropology graduate student who went on to become, like Sir Arthur Keith, a distinguished neurosurgeon. He became eminently qualified to evaluate the Morotopithecus spine bone.
Ultimately, Dr. Filler came to agree with Sir Arthur Keith’s view. In all respects the lumbar vertebra of Morotopithecus was close enough to a human vertebra to strongly indicate it had full bipedal capacity at 21.6 million years ago.
Dr. Filler went on to propose that other known forms of Miocene apes — namely Oreopithecus and Pierolapithecus — were also dominantly bipedal walkers, leaving the strong suggestion that if more fossils from the Miocene were available, more bipeds would be among them.
Dr. Filler (at UprightApe.net) presented a great deal of scientific detail in The Upright Ape: A New Origin of the Species, New Page Books, 2007. The same holds true for an easy-to-grasp article he wrote for Anthropology.net entitled: A Human Ancestor for the Apes?
Not worth reading is a long formal paper about the same material, found at www.plosone.org. Without Dr. Filler’s readable “translation” at Anthropology.net, the title tells us what we’re in for: Homoeotic Evolution in the Mammalia: Diversification of Therian Axial Seriation and the Morphogenetic Basis of Human Origins.
This quote is the paper’s summary conclusion:
“Clusters of homeotic transformations mark the emergence point of mammals in the Triassic and the radiation of therians in the Cretaceous. A cluster of homeotic changes in the Miocene hominoid Morotopithecus that are still seen in humans supports (the) establishment of a new “hominiform” clade and suggests a homeotic origin for the human upright body plan.”
That style is why ordinary people are so easily cowed into silent obedience to the obfuscating “High Priests” of mainstream science. Like all High Priests throughout human history, they isolate themselves from the “great unwashed” by what amounts to secret incantations that can be grasped only by those ritually trained in it.
Let me also say that Dr. Filler’s book is written for general audiences, though it, too, shows his scientific pedigree. I do recommend it with this caveat: He’s a credentialed mainstream scientist who solves problems by trying to conform them to the concept of evolution. However, his is not the gradual evolution of the strict Darwinists.
Like those who created Punctuated Equilibrium to acknowledge life on Earth clearly appears in periodic bursts, Dr. Filler is among the group of scientists that would replace gradualism with modularity theory, or Modular Evolution.
Again, modularity theory notes that bodies of all living things are comprised of distinctive parts. Like the different parts of automobiles, we have heads, torsos, upper and lower limbs, and organs to make us operate as we should.
Rather than trying to explain how the head of an ape gradually turns into the head of a human, modularity theory suggests that it happens quite rapidly, with extensive suites of genes mutating rather than random mutations in single genes.
This is “modern” evolution, lacking any need to worry about those pesky “intermediate” phases. So, once again, “It just … happens!” And once again, we find ourselves confronted with “God-worthy” miracles to explain what is becoming a new tenet of mainstream evolutionary science.
*****
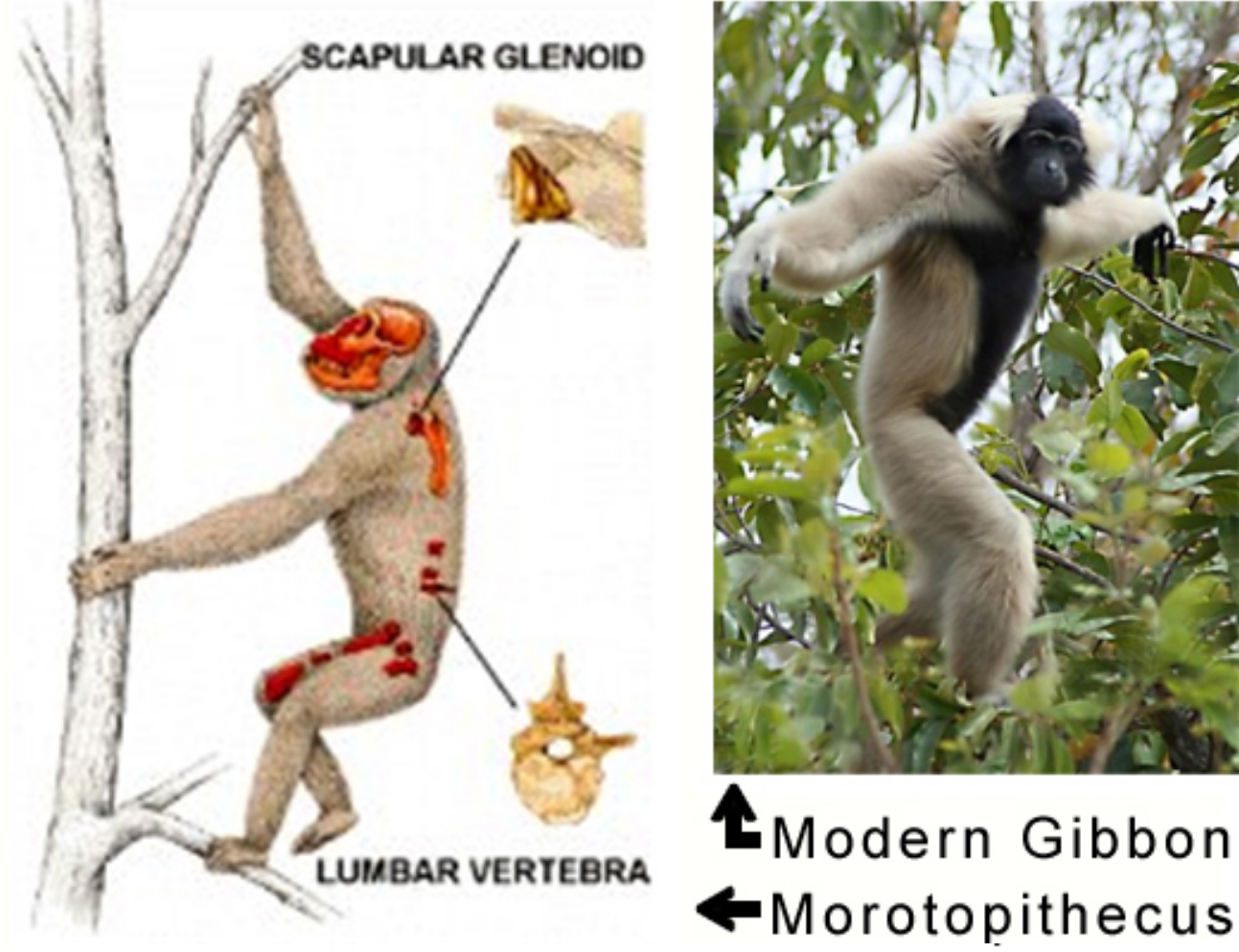 |
Getting back to Sir Arthur Keith and Dr. Aaron Filler, please understand that neither man ever suggested that any of the early Miocene apes, like Morotopithecus and the other potentially bipedal specimens, were anything other than 100% apes in all ways other than locomotion.
In the illustration at right we are shown how the mainstream explains the very unusual bones of Morotopithecus (highlighted in red), which they make physically look much like a gibbon. They contend that those extraordinarily variant bones are mere “adaptations” for “vertical climbing.”
Vertical climbing is indeed part of an arboreal lifestyle, with every monkey and ape doing it routinely. But what would require any ape like Morotopithecus to begin doing it so often that a wide array of body bones must be transformed to support a radically altered behavior pattern?
Nothing. It’s just the best excuse they have to make it seem as if all apes — even those whose bodies are obviously designed to be bipedal — remained dominantly arboreal, living in trees.
Why is that so important? Why can’t bipedal apes be living down on the ground at or near the start of the Miocene? Because no bipedal apes can exist until after chimps and humans split from their presumed common ancestor!
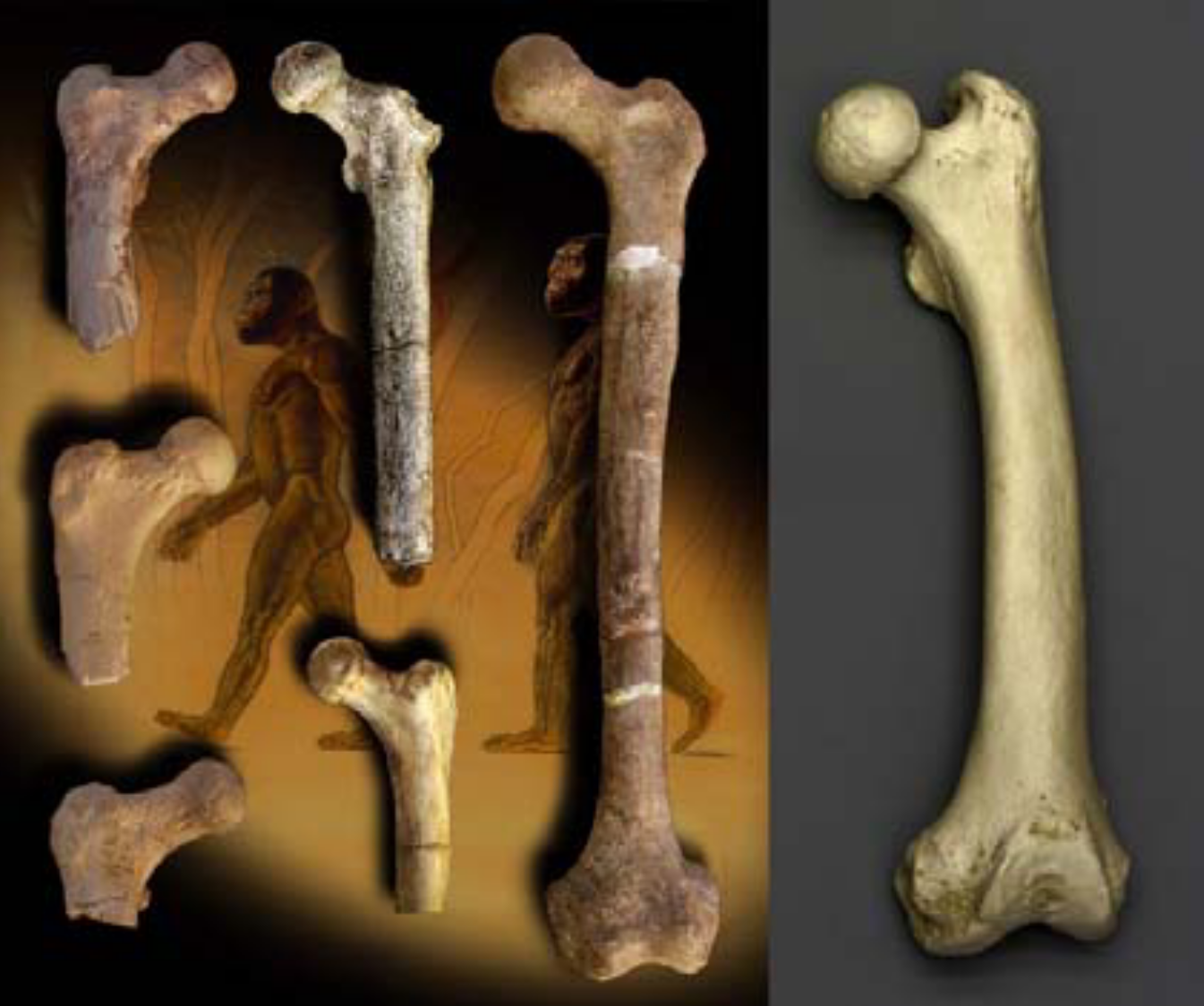 |
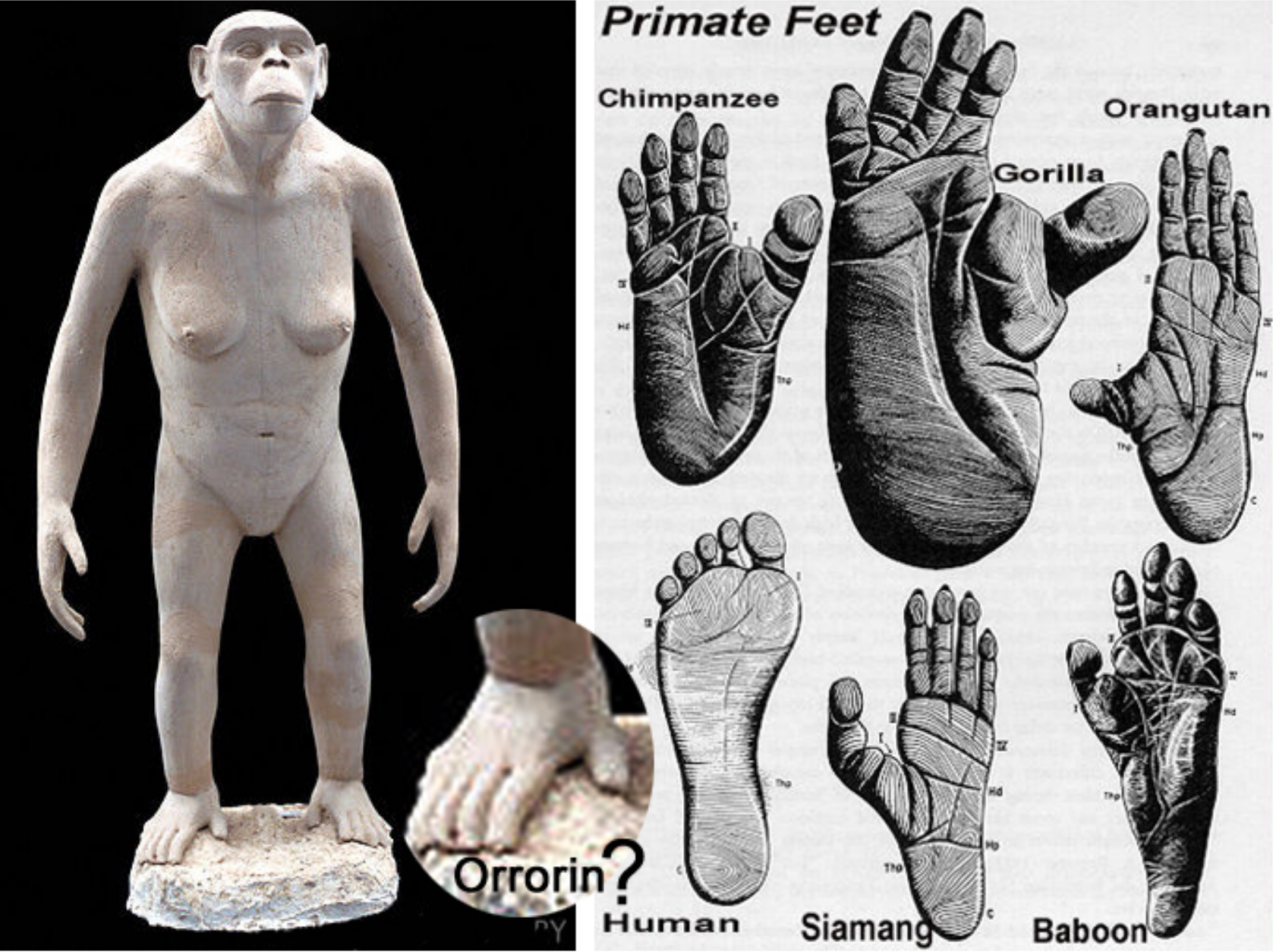 |
It depends on whom we ask. Geneticists know humans and chimps share 97% to 98% of the same DNA. Gorillas share 95%. This is taken as proof that humans and higher primates had to share a common ancestor at 5.0 to 8.0 mya. Anthropologists have a different perspective.
They keep discovering so-called pre-humans deeper and deeper in the past. These were apes in every way except they were bipedal, and the earliest candidate so far, Orrorin Tugenensis, dates at 5.8 to 6.1 mya. This is not enough time to “split” and then “evolve” so divergently.
In the photo at top right, Orrorin’s upper thigh bone (center, gray) is compared to early pre-humans (left & bottom), an early human (right), and a Bonobo chimp (far right). With such physical similarity to the bipeds, that is how it walked.
Unfortunately, no foot bones were recovered to show for certain that it had an opposed big toe.
In the depiction bottom right, notice that the length from shoulder to wrist equals from hip to ankle. This clearly makes Orrorin one of the “short-armed” bipedal apes from the Miocene epoch.
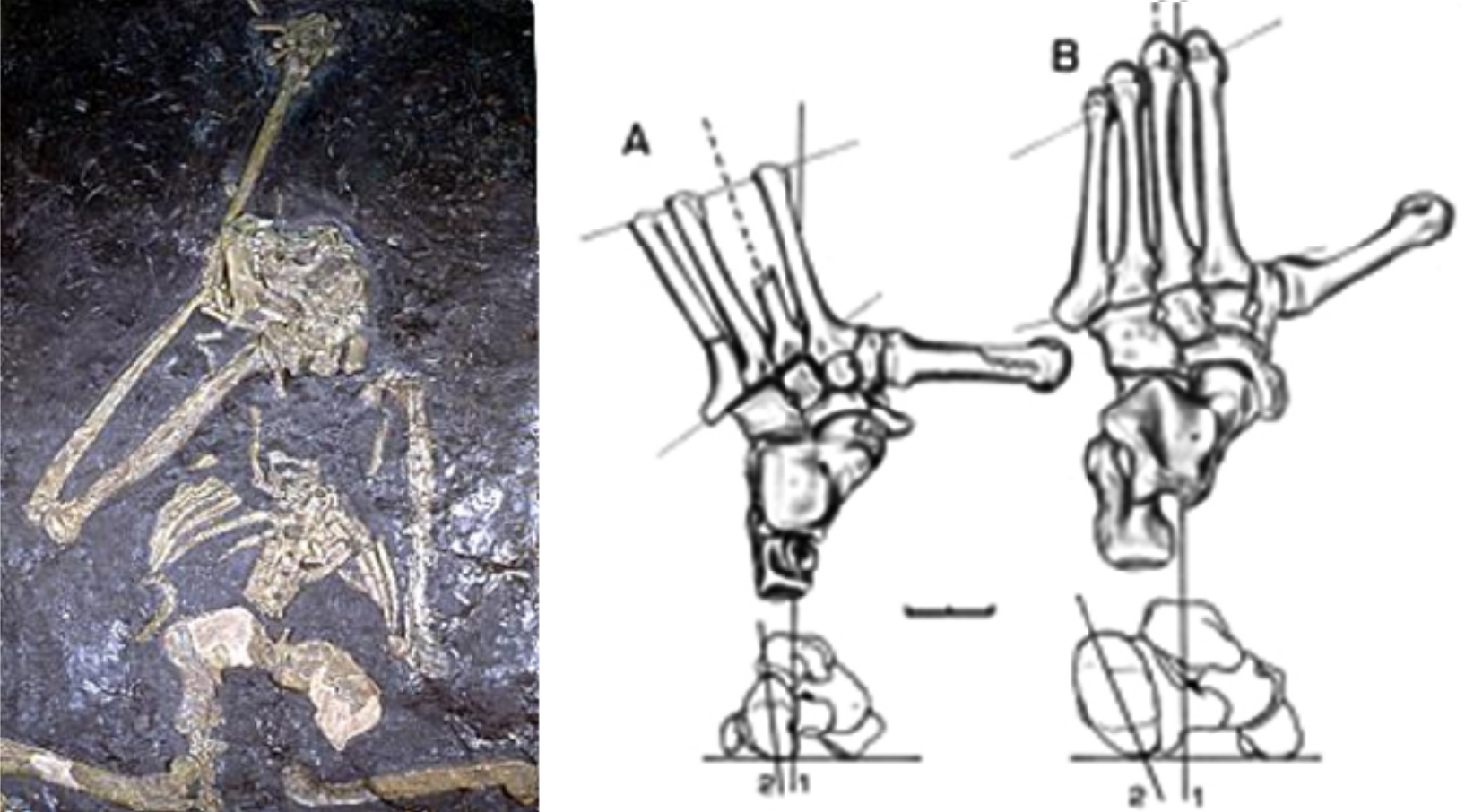 |
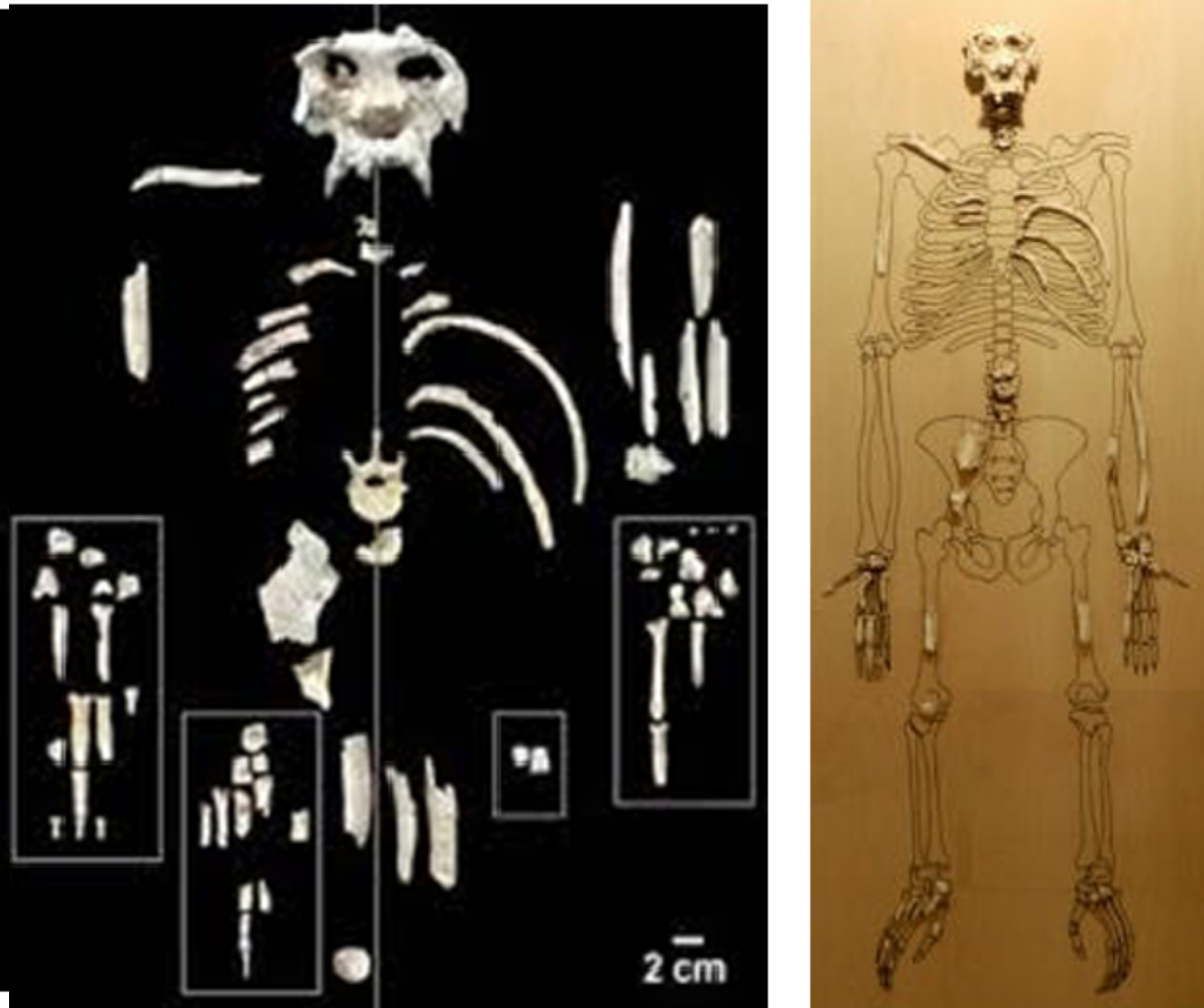 |
A serious conundrum faced by all mainstream apologists is that if bipedal apes existed at least 6 mya, it becomes a hop, skip and jump back to Oreopithecus at 8 mya. The “cookie monster’s” foot was clearly not human (“A” compared to a chimp foot “B” below). But its tripod structure made for a well-balanced stance when upright.
Also, notice in the photo on the left that Oreo had extra-long arms relative to humans, but not as long as chimps or gorillas. It appears to have been yet another mid-range, short-armed ape.
From Oreo at 8 mya, another 5 million years brings us to Pierolapithecus (bottom right) at 13 mya.
In the diagram at right we see the estimate of Piero’s arm length based on available bones. Again, they are longer than humans but not as long as chimps or gorillas. Another supposedly stooped-over, knuckle-dragging Miocene ape.
Also notable about Piero is that its feet also seem to sport opposed big toes, along the line of Oreo but not nearly as dramatic. And its foot is older by 5 million years! So the “better” foot clearly seems to belong to the “older” species.
 |
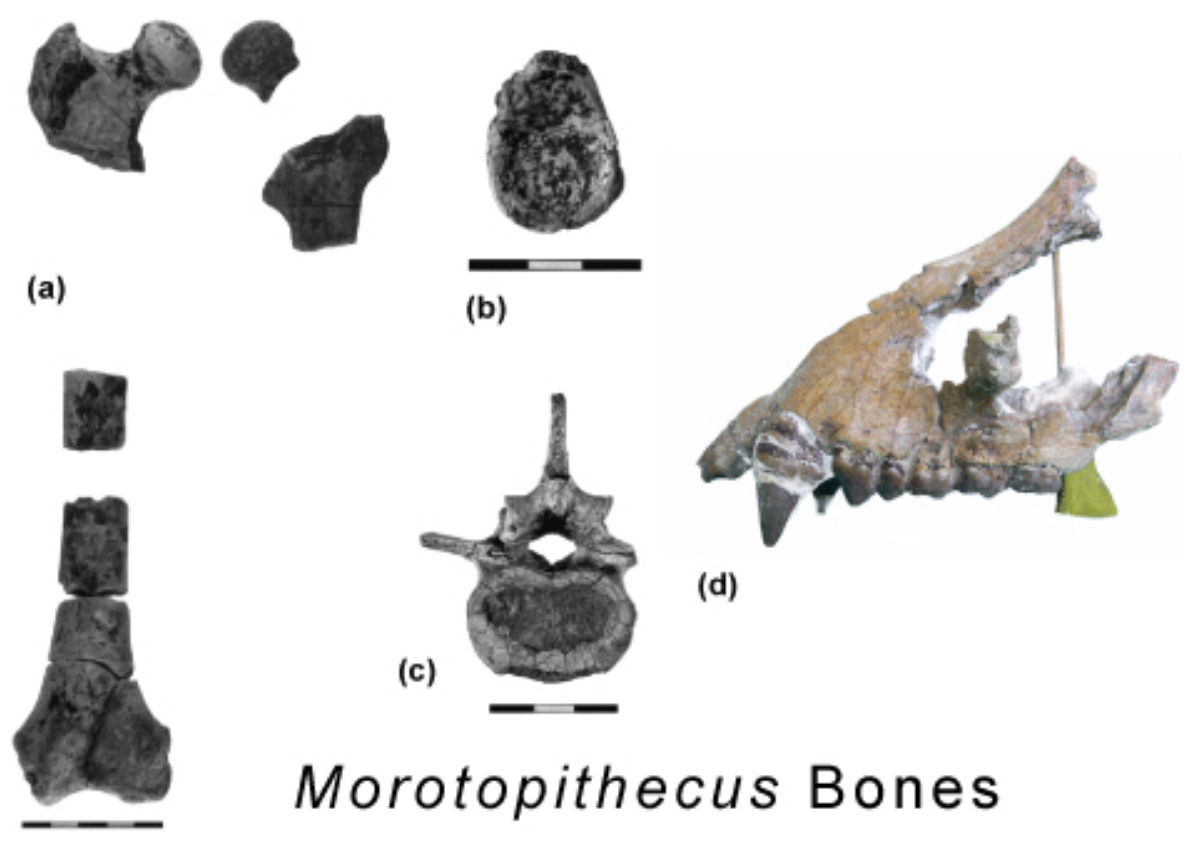 |
Another issue is that the right foot (on the left) has the most recovered bone fragments, including parts of the big toe. Yet it is depicted as much less opposed than the other toe. Why? Because the more the artist makes the feet seem primate-like rather than human-like, the more it subliminally supports the theory of evolution.
From Piero at 13 mya, another 8 million years brings us to Morotopithecus at 21 mya.
While not enough arm bones were recovered to provide an accurate estimate of length, Piero had a spine bone that was much like Moro’s, making it a solid bet that Moro’s arms were leg-length and it was yet another short-armed Miocene ape with hands dangling to its knees.
Now, with all of that said, it seems fair to ask why, if those three — Moro, Piero, and Oreo — can be counted as bipeds, why not others? Why not multiple others? Why couldn’t the Miocene begin with bipeds introduced first, as suggested by Sir Arthur Keith and Dr. Aaron Filler, along with several other well-qualified researchers?
Because to allow bipedal apes to exist during the Miocene era would contradict the idea that humans and chimps share a common ancestor, and that common ancestor is at the heart of the mainstream dogma that humans developed on Earth through a process of gradual evolution.
Is that dogma true? Are they right to insist that primates begat pre-humans who begat humans?
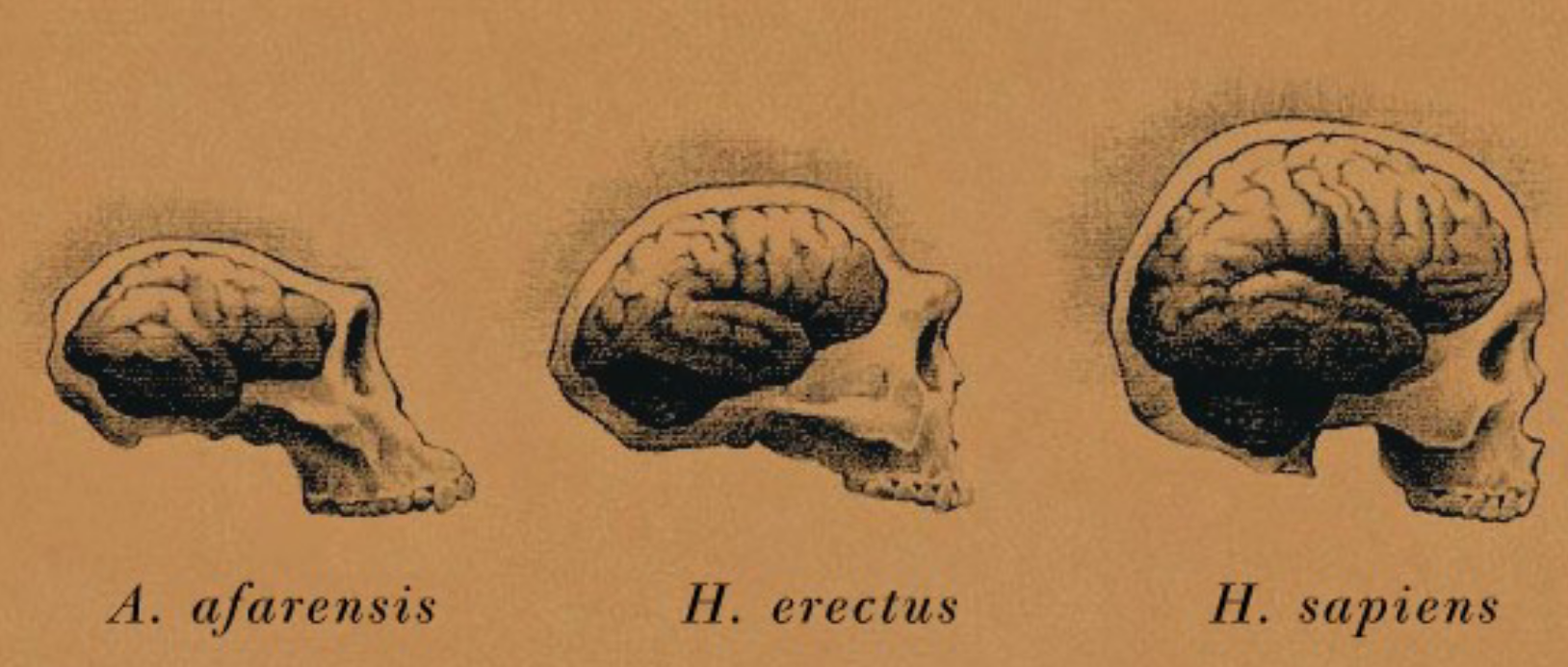 |
The “common ancestor” argument developed from Charles Darwin’s epiphanies in the middle 1800s. By the middle 1900s, it was ossified into rock-ribbed dogma that virtually all mainstream scientists would defend at all costs, in all ways.
Humans evolved from a common ancestor they shared with chimps at 5 to 8 million years ago. The proof science offers is a variety of upright walking primates referred to as pre-humans.
Despite strong conviction and strict adherence to their theory’s tenets, mainstreamers cannot make it correct. They can only guarantee that its adherents will be utterly blindsided by the truth when truth can no longer be suppressed.
This truth is that bipedal primates have been walking nearly everywhere on Earth since the start of the Miocene. They came with the first influx of tailless apes that appeared around 23 mya to establish dominance over the monkeys.
 |
Moro, Piero, and Oreo flow right into Orrorin Tugenensis (whom we met earlier) from 6 mya, at which point mainstreamers start calling any of the bipedal ancient primates “pre”-humans.
In fact, these creatures are “pre” nothing. They are “post” primates like Moro, Piero, and Oreo, and very likely many other such fossils that will be discovered in the fullness of time. However, for now we are stuck calling them pre-humans.
Following Orrorin at 6.0 mya is Ardipithecus Ramidus at 4.4 mya; then is Australopithecus Anamensis, 4.2 to 3.9 mya; A. Afarensis, 3.6 to 2.9 mya; Kenyanthropus Platyops, 3.5 to 3.3 mya; A. Africanus, 3 to 2 mya; A. Aethiopicus, 2.7 to 2.3 mya; A. Garhi, 2.5 mya; A. Boisei, 2.3 to 1.4 mya; A. Robustus, 1.8 to 1.5 mya.
Yes, I realize those convoluted names impede learning about this subject, but it is a deliberate part of “High Priest” functions to put what they know, or think they know, beyond the reach of ordinary people, so when they make mistakes, it becomes difficult to hold them accountable.
As we see in the early spelling bee at right, this has been a sensitive issue from our beginning.
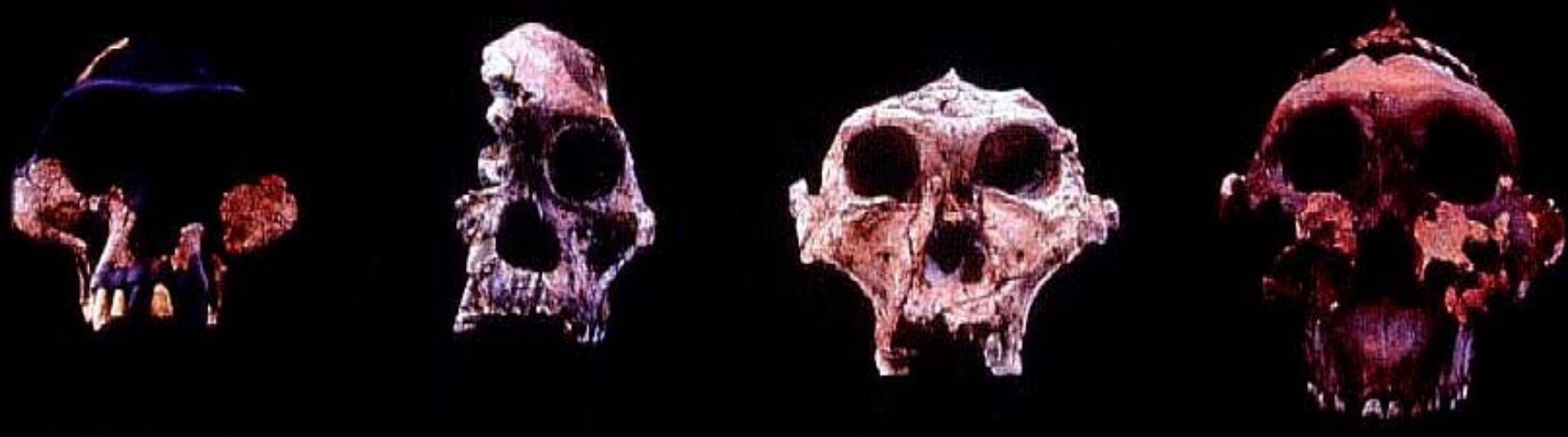 |
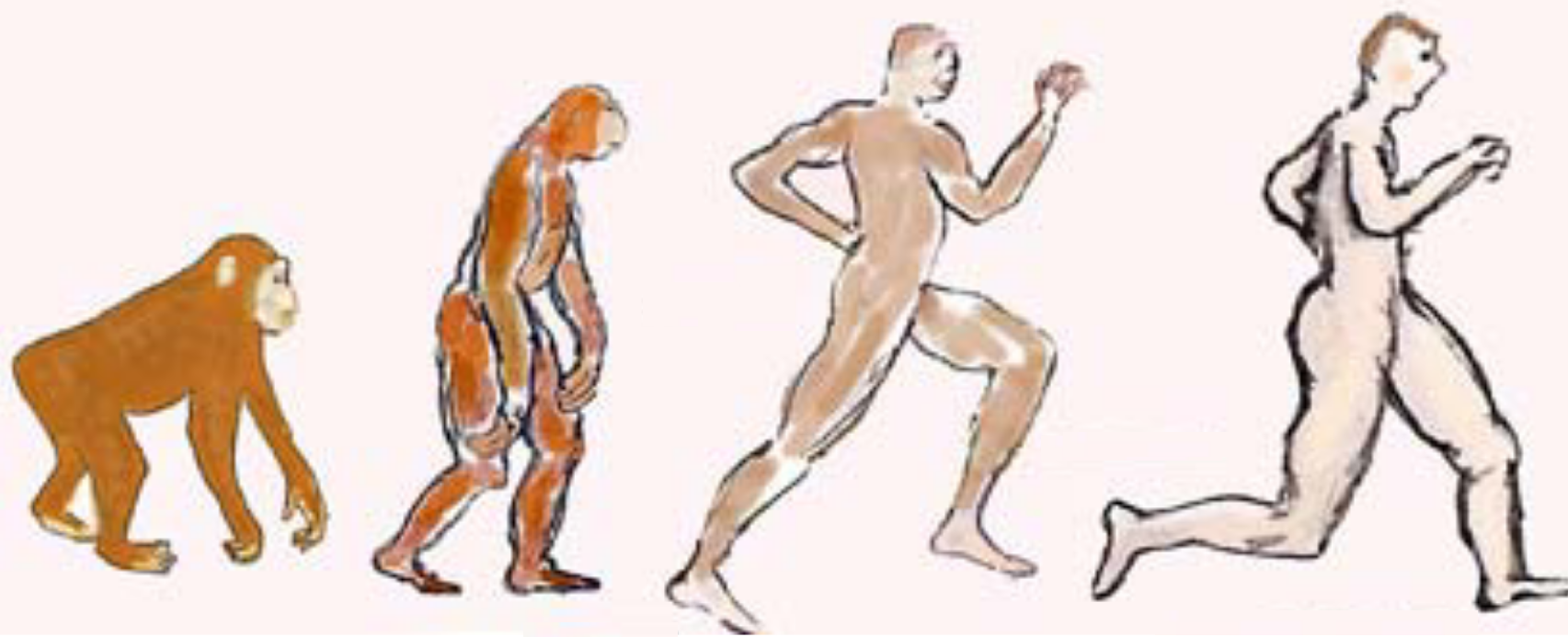 |
Australopithecines are basically a half-measure species — still fully covered with body hair like the primates they supposedly descended from, and still spending a great deal of time in trees, despite their physical capacity to be bipedal.
They appeared as two distinct types: (1) gracile; and (2) robust. The two dominant gracile types, Afarensis and Africanus (right top), were like bipedal chimps. The main robust pair, Robustus and Boisei (pair to the right side), were like bipedal gorillas.
Science preaches that at some point during the full flowering of the Australopithecine species, probably in the time frame from 3 to 2 mya, a “gradual transition” was made to full bipedality and to a loss of most of their thick pelts of hair.
 |
 |
 |
 |
How could that happen? How could two such fundamental aspects of their physiology be so thoroughly altered? To go from their presumed knuckle-dragging posture to erect? To go from a typical primate’s thick pelt of head-to-toe hair to a body with a greatly reduced cover of hair?
The way science tells it, the Australopithecines endured millennia of hot weather that dried out their forest homes, forcing them to the ground for periods that were more and more extended as they ranged farther and farther from trees.
On the savannas of Africa, they had to spend more and more time on their feet, which (as the mainstream scenario assures us) became better and better adapted for walking and/or running.
As they ran, their bodies overheated and had to sweat to dissipate that heat, so their body hair was in the way and, naturally, evolution was in some fashion directed to gradually diminish it.
Running was so critical to them because out on the savannas they encountered deadly big cats.
Faced with that problem, one might assume the Australopithecines would assess their situation rationally: (1) Ineffective weapons; (2) Lacking a tough hide; (3) No fangs or claws; (4) Awful foot speed; (5) Quite a few bricks shy of a load.
 |
With so many handicaps guaranteeing swift demise, you’d think it wouldn’t take long for Australo pre-humans to realize that out on the savannas they were hair-covered sitting ducks.
They were weak, helpless, slow-moving food bags; cake compared to mean African buffalos.
Does this make even a grain of sense? Maybe in an opium dream. However, lacking common sense has never stopped mainstream scientists from inventing a theory, then sticking with it.
The truth is that no matter how badly any forest home shrunk, the animals habituated to nothing but that would have to stay with it. They could not survive in a radically different environment, certainly not as different as forest and savanna.
No force is strong enough to compel any forest dwellers with few natural predators to venture en masse onto savannas teeming with big cats. This is merely another straw that mainstream scientists grasp in their desperation to support their precious, imperiled theory of evolution.
 |
 |
Homos. Yes, that word has multiple meanings that we all know have bemused many immature students, but this is science at work. Latin is the technical language of both science and religion because it is so difficult for ordinary people to understand, and in Latin, homo means man.
Homos appear in the fossil record 2.5 mya as a new kind of upright primate. Supposedly, they were a “transition” from the primitive Australo forms, but even a cursory examination of skulls shows that in all groups the change was more of a transformation. For the Boisei robust group (at right) the change was extensive (image on right).
The mainstream agrees that Australopithecines are rather primitive compared to the Homos. No argument there. Their problem is with plausibly explaining how such a huge physical leap ahead could occur with no intermediate transitions.
For evolution to rightly explain the appearance of humans, certain of Darwin’s “intermediate species” must be positioned between radically different groups. Australos and Homos require at least one, but thus far none have been found.
Even in the Australo group (bottom right), a wide range of differences exist between the robust kinds (top three and one at left), and the less primitive kinds (three at bottom right).
[The famous “Taung Child,” at bottom right, the first “gracile” type discovered, was a young A. Africanus found by Raymond Dart in 1924.]
Gradual evolution was not happening from 5 to 3 mya. For those 2 million years — at least — two very different types of ape bipeds, with multiple species of each type, existed cheek-by-jowl in apparent stasis in African forests and jungles.
Each species could easily represent descendants of bipedal apes that lived deep in the Miocene. Each could have direct ancestors that came with an initial group of “stock species” delivered to Earth 23 mya by Intragalactic Terraformers.
Bipeds like Moro, Piero, Oreo, and very likely others, spawned ancestors that are found in later geological strata, and because they are bipedal, science insists that they must be pre-humans.
They are not pre-human, they are post-Miocene bipedal apes, which when verified will provide convincing support for Intervention Theory.
***
 |
 |
Australopithecus Sediba, a relatively new find made in South Africa that dates to 1.9 mya, is a remarkably complete group of fossils that seem to have died together, or within the same time frame, and which provides some of the clearest views of ancient skeletons in the fossil record.
Many experts, most especially its discoverers (for the obvious economic reasons) contend that it best represents a genuine transitional species between the Australopithecines and the Homos.
The images (top right) represent a male child on the left and adult female on the right, found in such close proximity that they might well have been mother and son. Notice that, as usual with the so-called pre-humans, arm length is once again the same length as the leg, seen in presumably knuckle-dragging, short-armed Miocene apes.
These bipeds do have a unique blend of traits that seem a part of Australo morphology, and others more like Homos. For example, Sediba heel bones are more primitive than Lucy’s, the famous A. Afarensis of 3.4 mya. Yet its hands appear surprisingly advanced, capable of fine degrees of finger movement and manipulation.
***
 |
 |
 |
Regardless of how Homos came to appear at 2 mya, they were undoubtedly a step up from the Australos. They were still vastly more primate than human, but were indeed more human-like.
Homo Habilis is the first of the Homo group, at 2.5 mya. However, it is so primitive in so many of its physical features, in all likelihood it is yet another Australopithecine that is misclassified.
What gave Habilis its “Homo” credentials was being found with very basic stone “tools,” like what chimps and gorillas create in zoos when given stones. They bang them together and create “tools” like those of Homo Habilis.
Stone tools move through four distinct stages: (a) Oldowan, seen at upper left; (b) Acheulean, lower left; (c) Mousterian, upper right; and (d) Aurignacian/Upper Paleolithic, lower right.
The Oldowan “culture” extends to 1.5 mya. The Acheulean then goes to 100,000 years ago. The Mousterian and Aurignacian are more recent.
“Early” humans were alive about 200,000 years ago. Thus, the only “tools” likely not made by them were the chipped cores of the Oldowan and bi-faced hand axes of the early Acheulean.
Mainstreamers insist that all stone tools were created by incipient humans … but were they?
The first undoubted Homos begin with Homo Erectus at 2.0 mya. A number of possible sub-species fall into this category, including Homo Ergaster, at roughly the same time, 2.0 mya.
The Erectus/Ergaster debate centers on which one originated in Africa and spread out to India, China, and Indonesia. Or, conversely, did one appear in Asia and then migrate into Africa?
The answer is not clear-cut, except to say they do not seem to be the same species. Ergaster is a bit less robust than Erectus, which leads many specialists to conclude that Ergasters spawned humans, while Erectus spawned Neanderthals.
It is important to note that this refinement only occurred after mainstream science was forced to recant its prior cherished dogma that humans must have evolved from Neanderthals. But let’s save that discussion for later in the eBook. For now, let’s stick with meeting the early Homos.
 |
 |
To offer readers accurate representations of the early bipedal primates, Australos or Homos, is futile because most artists tend to depict them as, shall we say, rather “unattractive” humans.
We easily see how different pre-human skulls are when compared to humans, how brutish and primate-looking, so the faces artists create must accurately reflect the brutish/primate qualities.
However, the bodies they put under those faces are nearly always the slender shapes of humans. This practice is meant to subtly “brainwash” the unwary into believing without question that the mainstream’s evolutionary gospel is 100% true.
See how these bodies lack pelts? If you Google Australopithecine “images,” you’ll find most of the bodies are sleek and human-like, with butt-ugly faces and bodies only provisionally hairy.
Also notice the arms and hands, which swing at mid-thigh as human hands do. This is inevitably fudged. All pre-humans up to Neanderthals — and possibly including them — have arms longer than human arms, usually as long as their legs.
This is compelling evidence that all pre-humans are actually “short-armed” bipedal apes of the Miocene, living on to the time of “acceptable” bipedality, which is after Orrorin at 6 mya.
Also notice the clever touch of depicting the Homo Erectus at the left standing with a well-honed spear in hand, in front of a burning fire.
This, of course, is to overtly suggest they were skilled weapon-makers and masters of fire, but no evidence establishes they had such abilities.
 |
This is all part of a deliberately designed “spin” to dupe people into thinking pre-humans were precisely that — humans-in-waiting, beings only a few physical tweaks away from full humanity.
I used to play football against guys who looked worse than the one on the left, and I went to a junior high school dance with a girl who could be the sister of the one on the right. And those are supposed to be images of Neanderthals!
This is, as the Brits say, “utter rubbish.” Or, as we Yanks say, “total B.S.” The truth is that all the pre-humans, even Homos, were as far from humans as chimps — except for their bipedality.
Another bit of spin put on the “official” story of human evolution is the sequence of dates. It has a remarkable “flow” to it that makes it appear to happen in the expected Darwinian sequence.
After Homo Ergaster and Erectus at 2.0 mya comes Homo Antecessor, whom experts claim appeared between Homo Ergaster and the next evolutionary step “up,” Homo Heidelbergensis.
H. Antecessor appears at 1.2 mya, and then H. Heidelbergensis follows at 600,000. These are followed by the Big Kahuna of all pre-humans, H Neanderthalensis, the Neanderthals, early forms of which appeared at 300,000 years ago.
It used to be that what mattered to mainstream scientists was proving how humans “evolved” from Neanderthals. Now what counts is how, and to what degree, we interacted with them.
 |
Everything we are taught about Neanderthals is part of a Hitleresque “Big Lie,” the kind that if told often enough, and with enough conviction, will become truth no matter how deceptive it is.
Seen here are two depictions of Neanderthals. On the left is the blatantly primitive depiction, with a hand-axe on display, a prey animal slain, and its skull ready for mounting on a cave wall.
In addition, extra-heavy body hair is depicted, though he is modestly covered with a loincloth despite no shred of evidence (no pun intended) that Neanderthals ever wore skins as clothes.
Contrast that menacing figure with the far more human depiction on the right. The face is much less ugly, the body is much more “buff” in the modern style, body hair is greatly diminished, and it is posed like Rodin’s famous “Thinker,” subtly suggesting human-like mental powers.
Again, Googling Images for Neanderthals will provide a wide range of depictions, though very few show them as they almost certainly were — apart from most being ugly enough to make the proverbial freight train take a dirt road. But, as seen earlier, some are depicted as eerily human!
The truth is that all pre-humans were extremely robust, similar to modern body builders. They looked like a hirsute Arnold Schwarzenegger in his prime more than the rather mildly bulked-up bodies on display in dioramas around the world.
 |
Some notable physical traits were, as stated, uniformly thick bones, (supposedly) shortened forearms, wider and rather splayed hip bones, shortened lower legs with flattened lower leg bones, and truly butt-ugly faces (see below).
The next images (lower left) provide a reasonable view of their physical shape, but not necessarily of their skin color. (Most are depicted with light skin, resulting from what may well be innate racist tendencies, while very few are shown as dark.)
Another issue to discuss is how much body hair Neanderthals did or did not have. From the time they appear at 300,000 years ago, they clearly stay in areas plagued by frigidly cold weather, living near, or very near, the edges of glaciers during Ice Age maximums and minimums.
 |
 |
With an ability to move anywhere, they stayed where it was coldest during the 300,000 years they can be tracked through the fossil record.
That’s a long time to live in frigid weather with no record of ever using animal skins as furred body coverings, which Homo Sapiens Sapiens (humans) first do at 30,000 to 40,000 years ago, when the Neanderthals supposedly go extinct.
How does mainstream science talk their way out of this conundrum? They steadfastly insist Neanderthals should be depicted as seen below, battling their freezing environment by wearing animal hides tied together, despite a total lack of evidence of any kind that indicates they did.
 |
Another example of mainstream “spin” on this story comes from the few Neanderthal burials that have been found. Mainstreamers use this to suggest Neanderthals are all but human, having respect for their dead and a sense of spirituality. But, it would be another “God-worthy” miracle for mainstreamers if it were proved to be true.
What is quietly ignored is that all of the burial sites come very late in Neanderthal history, not until they had lived alongside Early Modern Humans and Cro-Magnons for 150,000 years. [Early Modern Humans appear 200,000 years ago, and Cro-Magnons at 60,000 years ago.]
This leaves two realistic options for explaining Neanderthal burials:
- They witnessed other species burying their dead and imitated those practices (unlikely after 250,000 years of not doing it); or
- The Cro-Magnons found dead Neanderthals, or maybe killed them as a result of hostile encounters, and provided the burials.
These observations force us to consider two conflicting assumptions about Neanderthals, only one of which can ultimately be correct.
The mainstream assumption is that they were entirely human-like, and therefore had to be at least partially hairless. If they lived hairless in near- or sub-freezing temperatures year-round, this would mean they had to cover themselves to protect their bodies from the incessant cold.
How could they do that? By wearing animal hides crudely tied together. But it is difficult to imagine them mastering the numerous complex tasks required to cure hides, though this seems the only option for human-like Neanderthals.
On the other hand, if Neanderthals were not as human-like as mainstream scientists want us to believe, and if they were in fact merely bipedal Miocene apes still living on as they always had lived, then they would be covered with a thick pelt of body hair that would keep them warm during and throughout any glacial weather.
 |
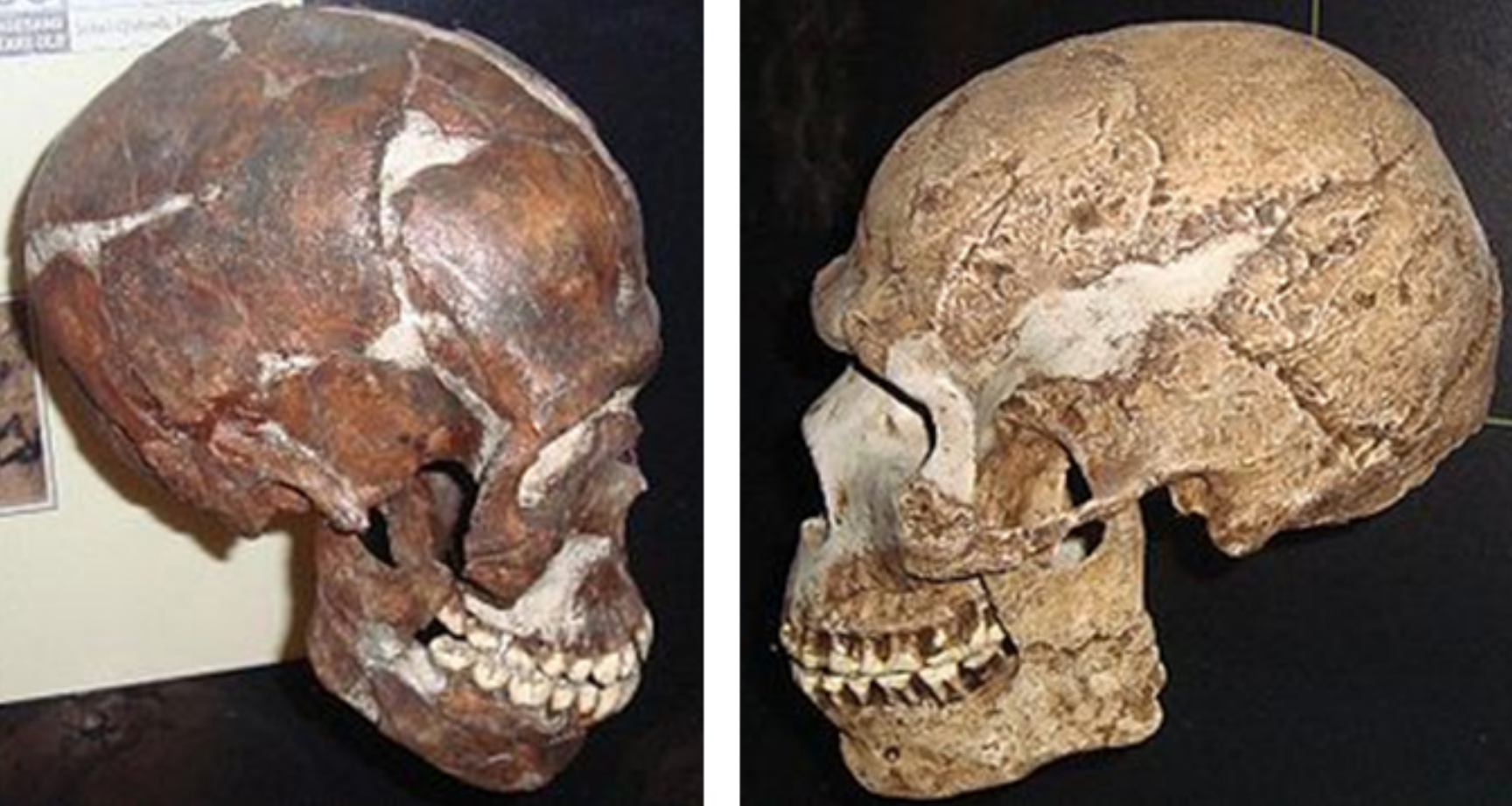 |
Next in line on the evolutionary ladder toward humanity is a recently mentioned group called Early Modern Humans (EMH). These appear in Africa around 200,000 years ago, exhibiting a mixture of “primitive” and “modern” traits, with some shading more of one way than the other. The consensus is that they are “human.”
Top right is the comparison of a modern human skull at the left and a Neanderthal in the center, with an overlay of both at the right. Notice the Neanderthal’s thick brow ridges and its heavy cheekbones, its weak chin, and the bulge at the rear of the head, known as an occipital bun.
These are very different species in every aspect of their skulls. Among higher primates they are the physical equivalent of apples and oranges. Now, imagine a physical blending of those two.
How would it look? Perhaps like the two skulls seen at bottom right, EMH species from around 100,000 years ago found in different caves located near each other in Israel — Quafzeh and Skhul.
The female on the left shows browridges much reduced from the male at the right, and her chin is much less than his, but the shapes of the rears of their heads are distinctly human, lacking any aspect of the Neanderthal’s “occipital bun.”
 |
 |
 |
These “types” of EMH supposedly evolved into the next step toward actual humans — the Cro-Magnons, living 50,000 to 60,000 years ago.
Cro-Magnons were not quite humans as we are today because, on average, they were larger and more robust. Most would fit well within a team photo of a typical football or rugby squad. But no one would mistake them for a Neanderthal.
For decades, mainstream science insisted the EMH species and Cro-Magnons had to evolve from Neanderthals because evolution was a gradual, step-by-step process. That meant any more “sophisticated” species had to follow the Neanderthals in the march to full humanity.
Various forms of the iconic image above have been imprinted on the brains of students around the world. Today, most people accept as gospel that pre-humans smoothly “transitioned” from knuckle-dragging to modern bipedality, while at the same time their arms shortened and body hair receded, with hardly a bump in the road.
This concept of orderly marching ignored the fact that only 100,000 years separated the first Neanderthals, at 300,000 years ago, from the first EMH types at 200,000 years ago.
That was clearly not enough time for gradual evolution from one to the other to occur. Only punctuated equilibrium or modular evolution allowed for such rapid transformations. Also, those two lived in conjunction — and sometimes literally adjacent! — for at least 150,000 years.
At left (in the bottom right) is Skuhl cave in Israel where EMH are estimated to have lived from 120,000 to 80,000 years ago. On the right is the Kebara cave near Skuhl, where Neanderthals supposedly stayed from 60,000 to 50,000 years ago. But all such dates tend to be very rough approximations.
Regardless of the actual intervals both species might have lived in such caves, it is possible they traded occupation times as the Ice Ages waxed and waned, especially if, as assumed, EMH possessed significantly less body hair.
As glaciers extended south toward the Levant, the EMH groups would move south to warmer regions. Neanderthals would move in ahead of the ice and stay until it receded, following the cycle. And, of course, during times of overlap there might well have been sexual mingling.
Despite these and other gaping holes in their logic, mainstream science had to prop up the “marching” scenario because humans had to evolve from some prior species — one way or another. Thus, nearly all drawings, paintings, and sculptures in dioramas were designed to support the mainstream’s century-old dogma that Neanderthals somehow spawned humans.
***
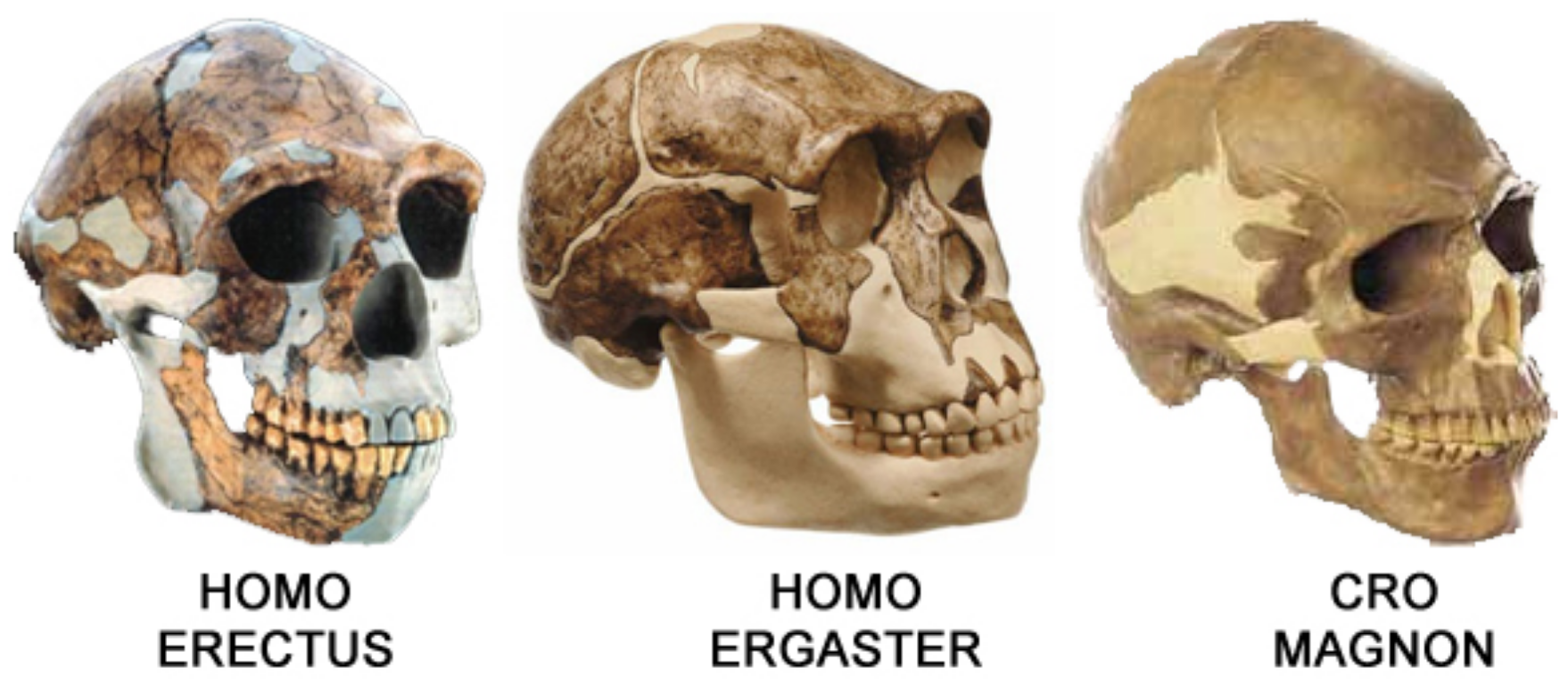 |
Everything went well for mainstreamers until, quite unexpectedly in 1996, geneticists made a disconcerting discovery: the Neanderthals and humans did not share enough mitochondrial DNA! If humans were directly descended from them, the comparison should have been closer.
Mainstreamers circled the wagons around their evolutionary dogma. Upon losing Neanderthals as our direct ancestor, they shifted their focus to H. Erectus and H. Ergaster as the candidates most likely to evolve into EMH and then Cro-Magnons. Both had much more time to evolve.
[The illustration at right shows the two options mentioned above — that either Homo Erectus or Homo Ergaster evolved into Cro-Magnon.]
The advantage of working with fragments and pieces of fossils is that they are so incredibly subjective. One day they can mean one thing, another day they can mean another thing. The bones don’t change, only the agendas of those “interpreting” them to suit a favored ideology.
Each fresh discovery of major consequence — like the recent Sediba find — changes the game, and the game plan, for mainstreamers. But they always find a way to make every piece of new information become part of the glue that holds together their theory of human evolution.
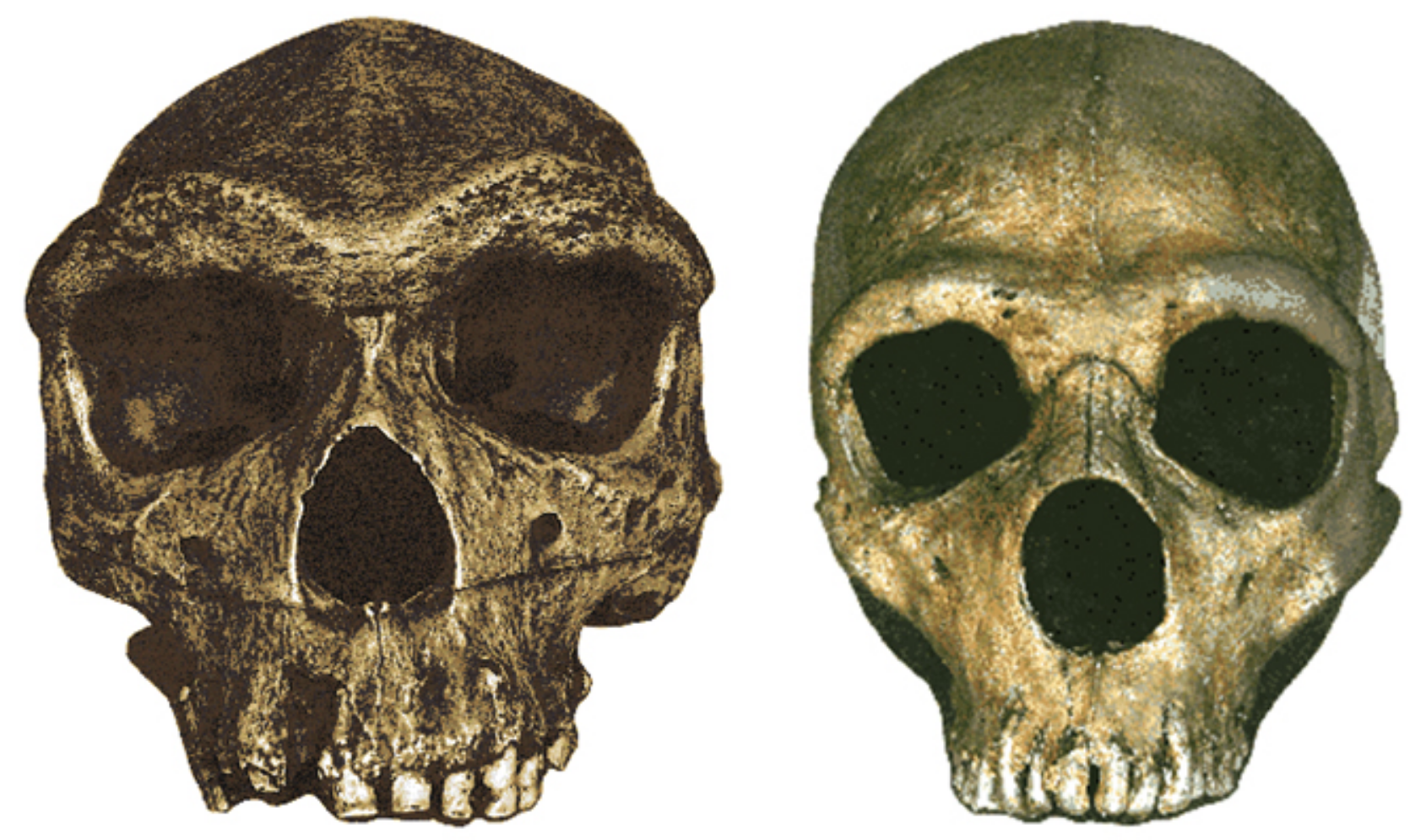 |
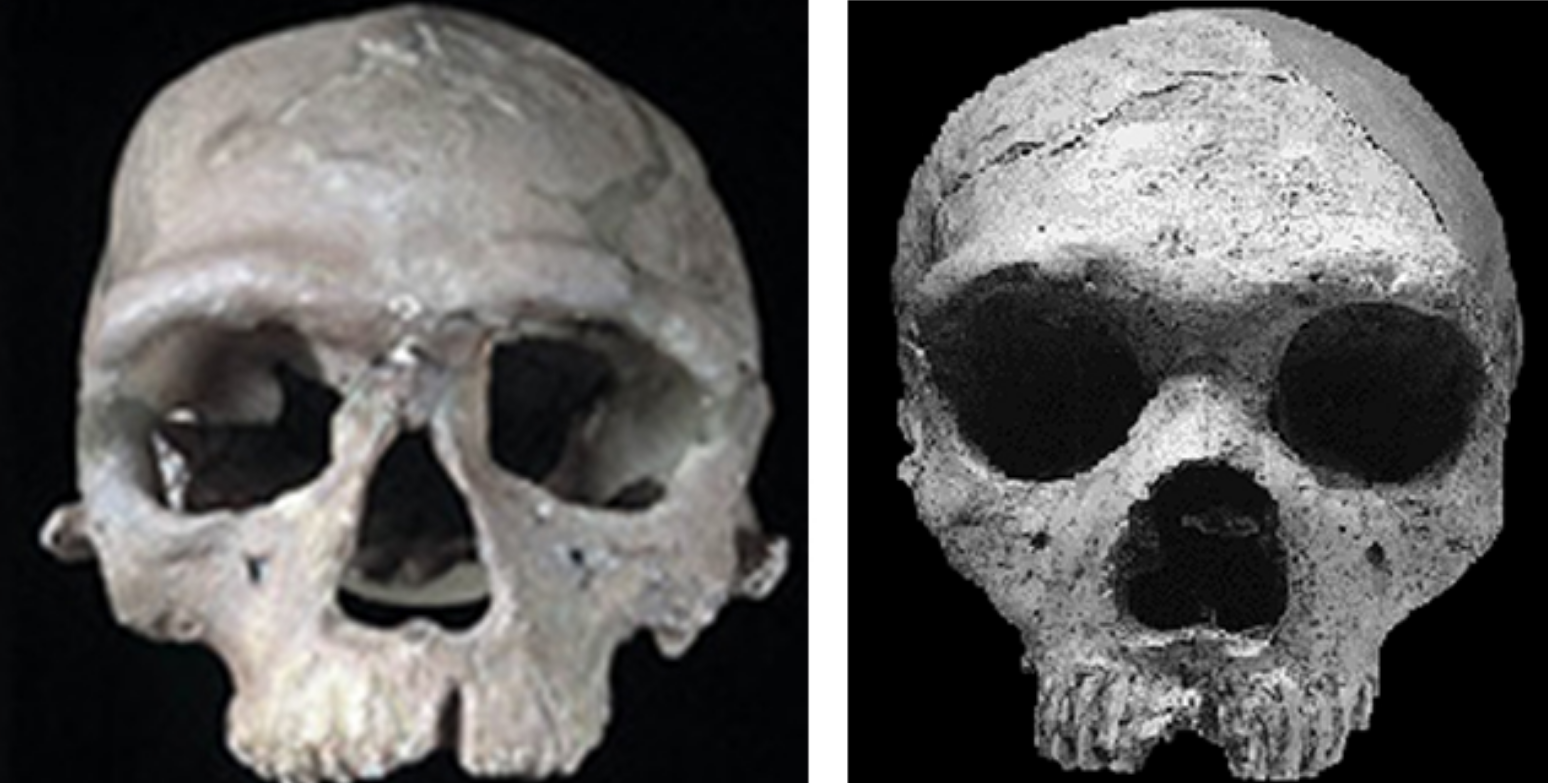 |
 |
Regardless of what led to the Cro-Magnons, they were exceptionally different from all that came before them, and there is no doubt they are the direct biological ancestors of humans.
Let’s begin with a review. In the top image, on the left is Homo Heidelbergensis from 600,000 to 400,000 years ago. On the right is a Neanderthal from a cave at Shanidar, Iraq, around 80,000 years ago.
Those skulls were owned by a couple of very rough looking individuals, whoever they were. But, it is reasonable to assume one could have microevolved from the other in 300,000 years.
Now, here is a challenge. In the middle image, one of the skulls is a Neanderthal and one is an Early Modern Human (EMH). Which is which?
If you’re having trouble, check the nose areas. Neanderthal noses are very wide and splayed.
As seen above, it is a much bigger jump from Neanderthals to Cro-Magnons, distinctive with rectangular eye sockets, reduced brow ridges, high foreheads, narrow uplifted noses, reduced cheekbones, and jutting chins. Microevolution seems much less likely to have produced this.
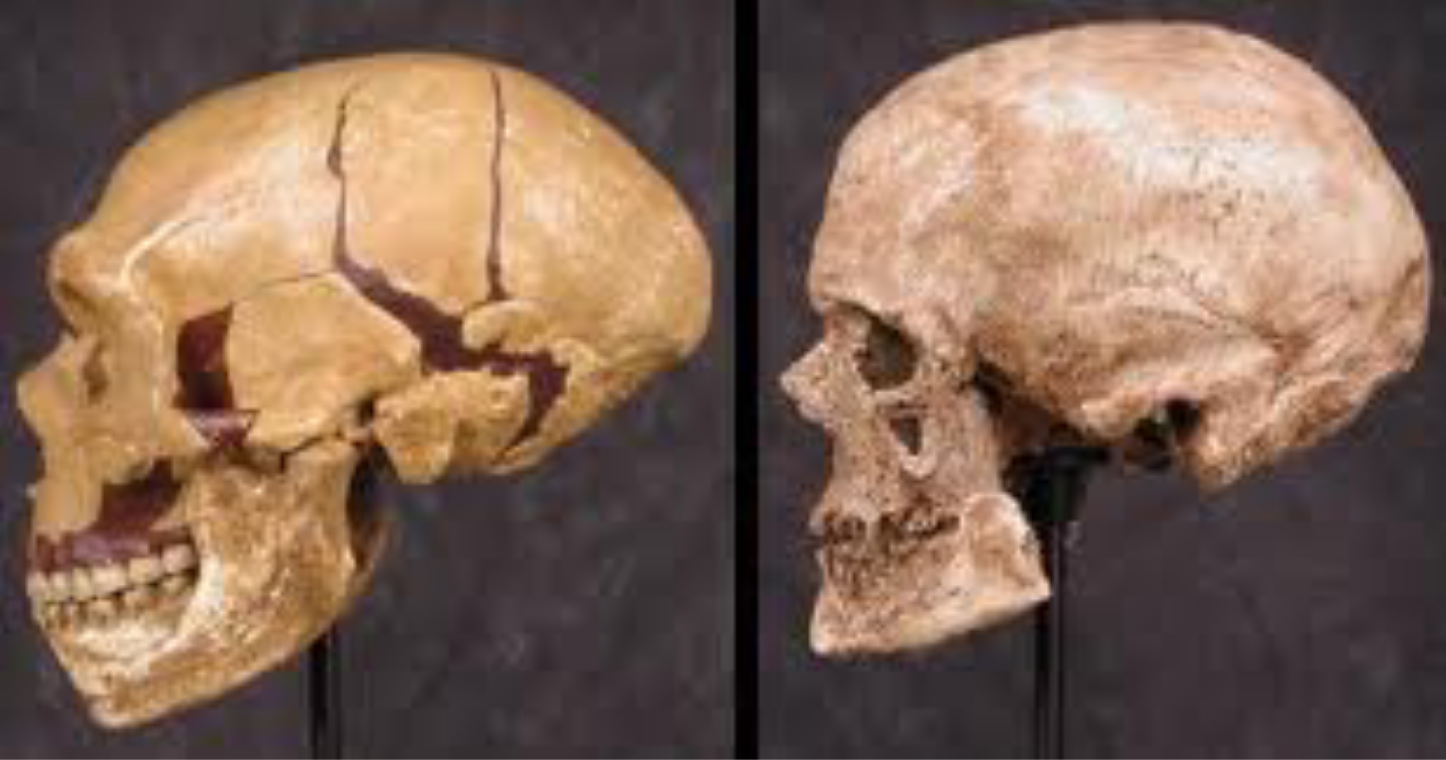 |
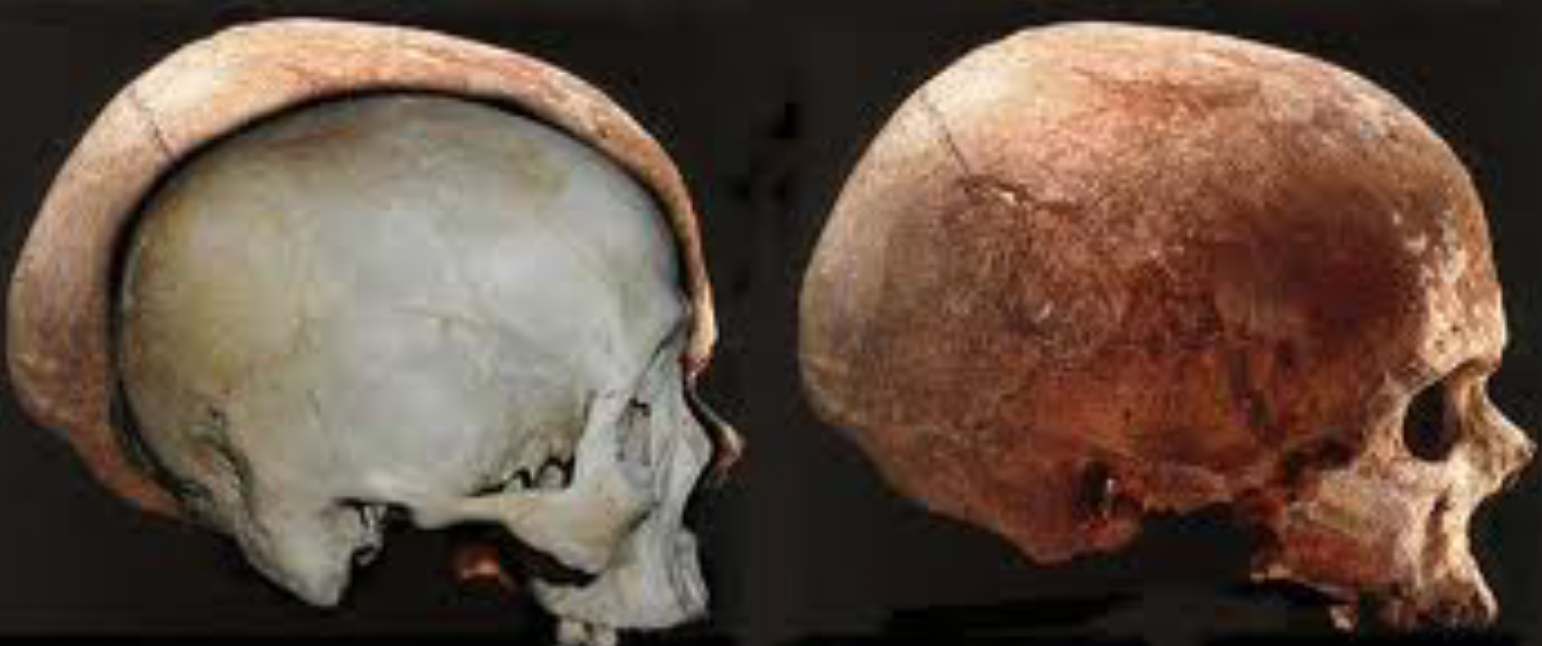 |
Here is another comparison, this time in profile, of Neanderthals and Cro-Magnons. Again, it is difficult to imagine microevolution creating so many physical changes in only 200,000 years.
For as human-like as Cro-Magnons were, they still differed significantly from us, including a somewhat larger brain (as seen in the bottom right image).
Despite these contradictions to classic evolution theory, mainstream science remains in a catbird seat, able to dictate how images are displayed so that they project the “correct” interpretation.
 |
 |
For example, the group at left are arranged to show a smooth transition from Australopithecus at bottom left, through the “transition” species, middle, and to a modern human at bottom right. When viewed from this perspective, it is little different from the “horse” chart we saw earlier.
If we listen to the “experts,” there is simply no argument, no doubts about it, no other choices.
As seen in full on the next page, a seemingly logical “flow” of all species has been arranged, and no matter how much they look like bipedal primates that walked out of the Miocene, they are considered to be only pre-humans — period.
Is that true? Not necessarily. Instead of humans macroevolving from Miocene apes, maybe the apes continued living as usual for 20+ million years, microevolving as circumstances dictated, while humans came to be here by other means. This idea is fully explored in the next chapters.
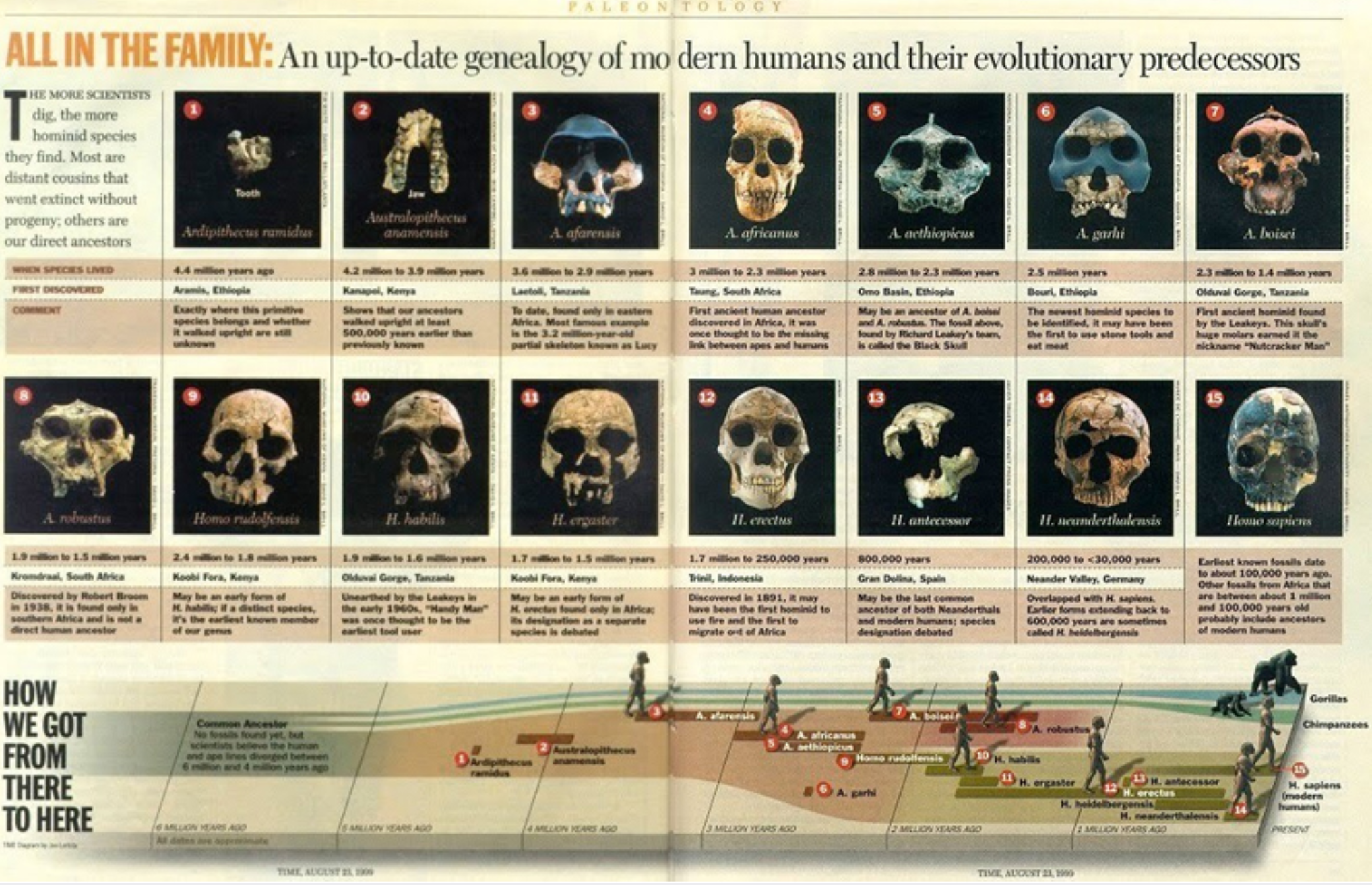 |
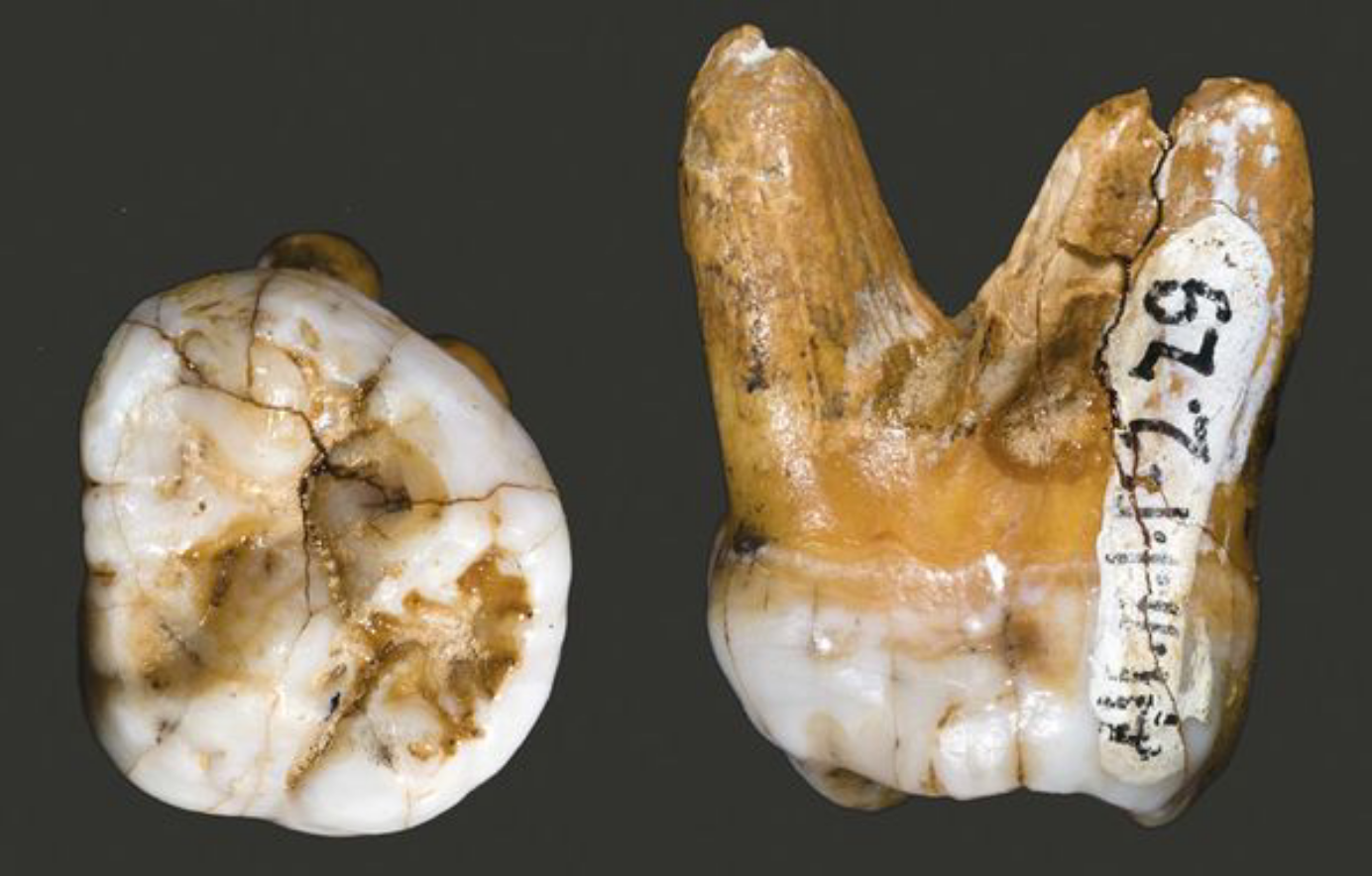 |
 |
Recall that in 1996, geneticists discovered that Neanderthals and humans did not share enough of their mitochondrial DNA (mtDNA) to have any kind of biological relationship between the two species. This was a huge jolt to mainstream anthropologists, who were certain that humans had to evolve directly from the Neanderthals.
Reluctantly, anthropologists shifted humanity’s closest predecessor to either H. Erectus or H. Ergaster (leaning toward Ergaster), but they clung to the belief that humans must still have some kind of biological link to Neanderthals.
Finally, in early 2010 the anthropologists were vindicated. Advanced genetic testing showed that non-African humans carry between 1% and 4% of Neanderthal DNA, while Africans carry none. This suggests interbreeding between them occurred in places where they lived adjacent to each other, in Europe and the Near East.
Advanced DNA analysis then led to an even more startling discovery — another species of late pre-human was found! Called Denisova, after the cave in Siberia where its bones were located, the new species is assumed to look like Neanderthals despite their genetic differences.
So far, the only physical remains found of the Denisovans are a finger bone and the tooth (seen top right), which is more robust than Neanderthal teeth. This corroborates the DNA findings.
A perplexing oddity that came from the DNA results is that while Neanderthals contribute 1% to 4% to humans outside Africa, the Denisovan species seems to have contributed up to 5% of the DNA genome of Melanesians — the people of New Guinea, Fiji and scattered islands in the western South Pacific (bottom right is a Melanesian).
A further puzzle is that in Europeans, Asians, and Melanesians, one segment of DNA from Neanderthals and Denisovans is known as the HLA histocompatibility complex. These are protein-encoding genes found on chromosome 6, which help protect humans against assaults by various bacterial infections and viruses.
How would something as specific and useful as that happen to find its way into the genomes of all non-Africans? The mainstream is calling it sheer blind luck that some of us had sex with some of them, and some genes we happened to gain on permanent loan from those encounters are a vital boost to our health. Another miracle!
***
 |
If we keep in mind Intervention Theory and the Intragalactic Terraformers, it becomes easy to suggest that maybe humans were brought here to Earth, or genetically created here, and those doing the bringing or creating understood that the new species would need a healthy dose of inoculation against Earthbound pathogens.
What would be the best way to do that? Well, one way would be to find a species genetically compatible with the new species, and transfer their localized pathogen-fighting ability to the new species. Of course, this requires a level of genetic understanding that is vastly beyond our current knowledge base, but if Terraformers are behind it, such things would be easy for them.
In any case, the Neanderthals and Denisovans just happened to pass along to all non-African humans — once again … as if by magic! — the specific genetic additions needed to fare better against the pathogens found outside of Africa.
As for Africans, the exclusion of such additions to their DNA might be explained by recalling that they are the oldest true humans (shown by mtDNA studies), so they might have received an earlier complex of pathogen-fighting DNA that was unique to the African environment.
Accepting that assumption leads us to suggest that later, when the Terraformers decided to expand the reach of humans into other places on Earth, they added what they thought those migrating groups would require while there, including, perhaps, variations of skin color to cope with variations in sunlight reception in order to properly absorb enough vitamin D.
Before we explore that radical suggestion any further, let’s examine the equally contentious question of whether hominoids exist or not.
 |
Technically, the term hominoid includes all members of the ape family — the great apes (chimps, gorillas, orangutans), the lesser apes (gibbons), and the in-name-only apes (humans).
However, since DNA testing proved the human and chimp genomes are 97% to 98% the same, mainstreamers have seized their opportunity to infuriate Creationists and Intelligent Designers by insisting that it means humans and chimps belong in the same biological grouping.
Scientists know that if chimps and humans are linked as closely as that, it greatly strengthens Darwin’s original argument for evolution from a “common ancestor.” So, mainstreamers have developed, and are diligently promoting, a new term for humans and chimps (and some want to include gorillas, who share as much as 95% of the same genomic package) — hominins.
That being so, I and others in the worldwide community of alternative researchers — most particularly in Russia — have usurped the no-longer-in-favor word hominoid to represent the numerous varieties of upright walking, hair-covered, ape-like creatures known on every continent (except Antarctica) by names like:
Bigfoot, Sasquatch, Abominable Snowman, Yeti, Yowie, Almas, Kaptar, Sedapa, Agogwe, Oh-Mah, Mapinguary, Wauk-Wauk, Hsueh-Jen, Meh-The, Gin-Sung, Apamandi, Teh-Lma, Toki-Mussi, Muhala, Dwendi, Kang-Mi, Orang Pendek, Golub Yavan, Jez-Termak, etc.
Let’s consider the pair at top as prototypes for the creatures listed above, and dozens of others. They fit the physical pattern of four groups of robustly built, hair-covered, bipedal primates.
| 1 | 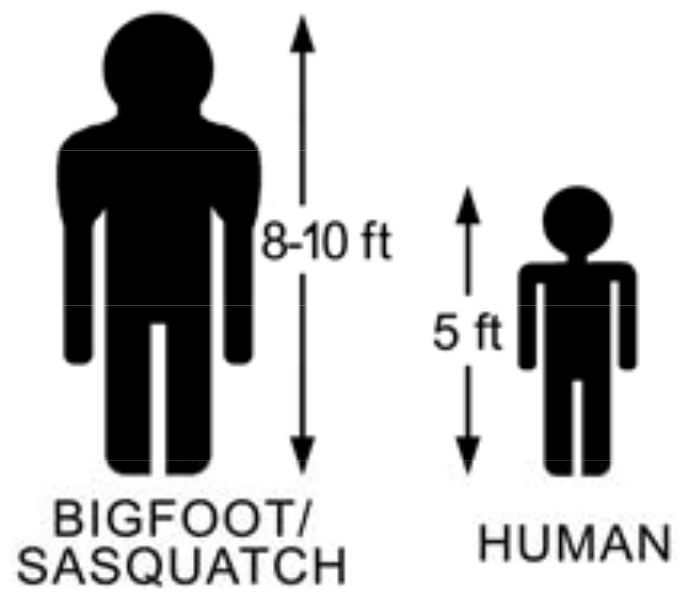 |
| 2 | 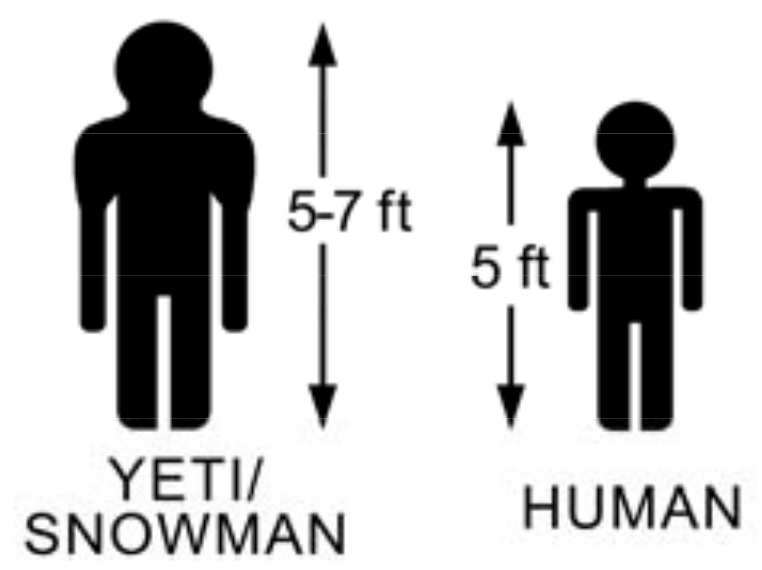 |
| 3 | 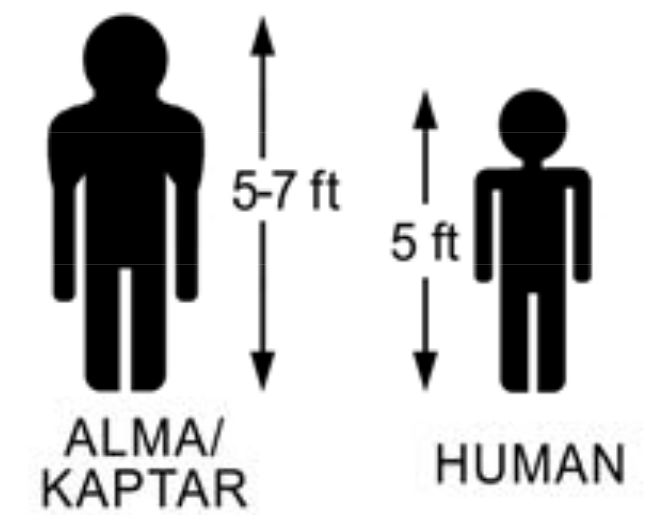 |
| 4 | 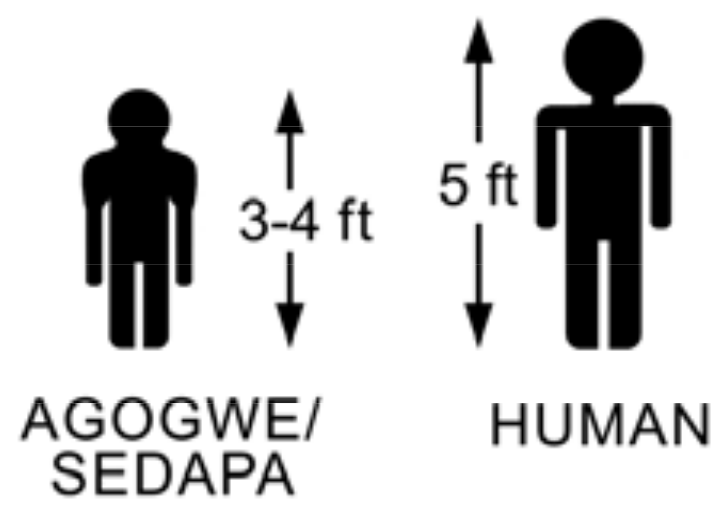 |
(1) The bigfoot/sasquatch type. These are the giants everyone is most familiar with. They are 8 to 10 feet tall and weigh 400 to 700 pounds. They live in thick montane forests that drape mountains on most continents of the world.
(2) The abominable snowman/yeti type. These are equally famous. They are man-sized, at 5 to 7 feet tall, weighing 300 to 500 pounds. They are the most primitive of the four types. They live exclusively in forested valleys and gorges within the five Himalayan mountain ranges, which together would cover the United States.
(3) The almas/kaptar type. These are basically unknown in the West because they dominate in eastern Europe, especially in lowland forests of Russia and several countries of the old Soviet Union. They, too, are man-sized, 5 to 7 feet tall, weighing 300 to 500 pounds. They can exhibit shrewdness, even cunning. Some researchers, including me, suspect these creatures may well be Neanderthals living on into modern times.
(4) The agogwe/sedapa type. These are the “pygmies” of the group, standing 3 to 4 feet tall, weighing 100 to 300 pounds, and living in the band of rain forests and jungles that girdle the Earth around its equator. These can exhibit cleverness, and may be the smartest of the four.
This type became world famous recently with the discovery of some of their bones in a cave on the island of Flores, in Indonesia. Dubbed “Hobbits” by the media, the creatures that left those bones were what Flores natives call ebu gogo, their name for the Orang Pendek, which is what they are known as in nearby Sumatra.
 |
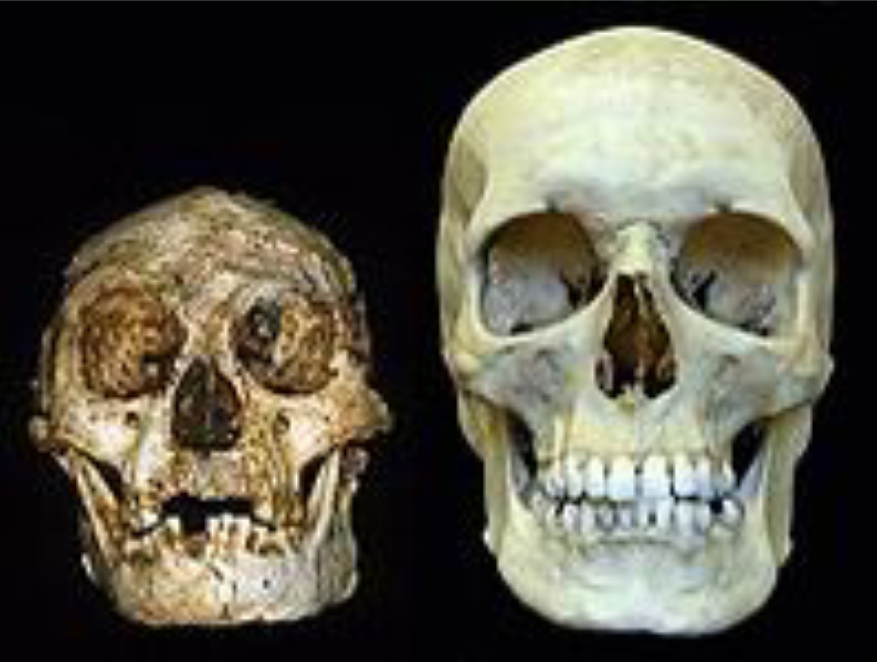 |
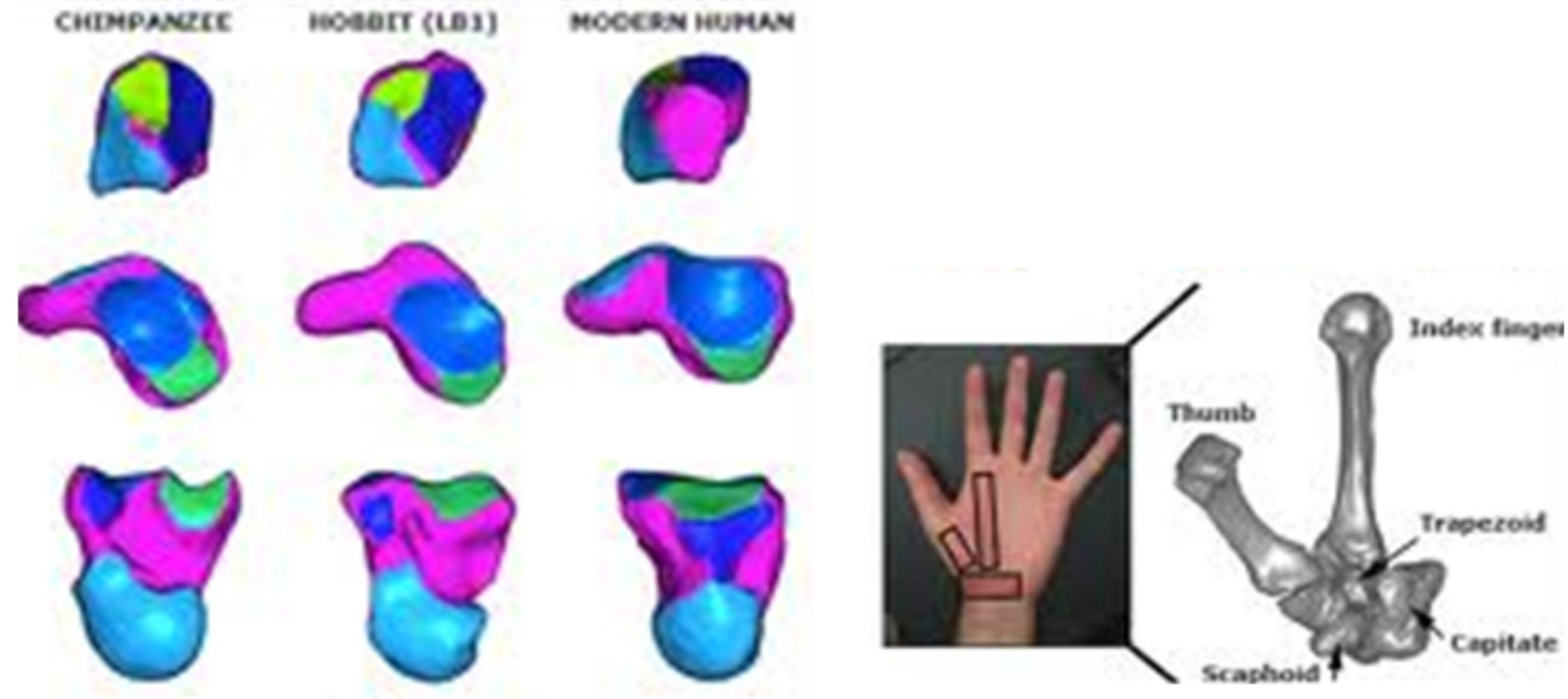 |
Top left is how Hobbits are depicted by the usual mainstream artists hired to sustain the “if it walked upright, then it has to be pre-human” dogma. Top right is how locals consistently describe these jungle dwellers, with muscular primate bodies and thick manes of head hair.
At least the artist of the “official” drawing was honest enough to paint the Hobbit’s extra-long arms as they would have been when they lived. Again, this is a commonly consistent trait of all pre-humans and of all bipedal Miocene apes. A strong link between them is glaringly obvious.
Nearly all mainstream scientists insist that the Hobbits must be pre-humans, despite one of the discoverers, Mike Morwood, admitting on the U.S. TV show, 60 Minutes, that no bones in the Hobbits closely resembled any human bones.
Among many examples of radical differences are, of course, the skull, which is clearly that of a primate; but also in bones as innocuous as the wrist, which are also more like a chimp than a human. At right, examples of both are compared.
The four hominoid categories listed above have resulted from thousands of eyewitness reports given over hundreds of years. While variations do exist within each group, and more than one species is in each group, categorizing them as having four distinctive body sizes seems valid.
The main point to understand is that hominoids are real, and they represent living Miocene apes that were bipedal as far back as the beginning of the Miocene epoch, 23 million years ago.
Naturally, mainstream scientists insist they can not be real, but not because they’re not actually real, but because they simply can’t be real if the dogma of human evolution is to have any hope of being believed by the majority of people.
 |
Science bases a blanket dismissal of hominoids on their insistence that no proof offered in their behalf can be acceptable because it would not conform to the current evolutionary paradigm.
Scientists have no proof hominoids do not exist. They merely stand on self-proclaimed authority and insist that whatever they declare is divinely right until they inform us otherwise. In the case of hominoids, that won’t happen until a body — dead or alive — gives them no choice about it.
However, that does not mean convincing proof has never been presented. In actuality, it has on many occasions. In this eBook, I can only focus on some of the best, but please trust that dozens of other topnotch cases exist in the literature.
In my opinion, the finest collection of evidence is found in Canadian Christopher L. Murphy’s tome, Know the Sasquatch/Bigfoot, published by Hancock House in 2010. It is a high-gloss coffee-table book filled with photographs, illustrations, and diagrams that present an overwhelming case for hominoid reality.
This book goes into great depth about two of the best examples of proof — hominoid tracks, and the world-famous Patterson-Gimlin film.
Tracks are the very best form of proof because they are so numerous and so easy to judge for veracity. Wherever they are found around the world, they invariably tell the same story.
Regardless of the body size of the creatures that make tracks — from tiny mice to elephants — the imprints they create contain certain elements in common. Now we will examine some of those elements to see what they can tell us.
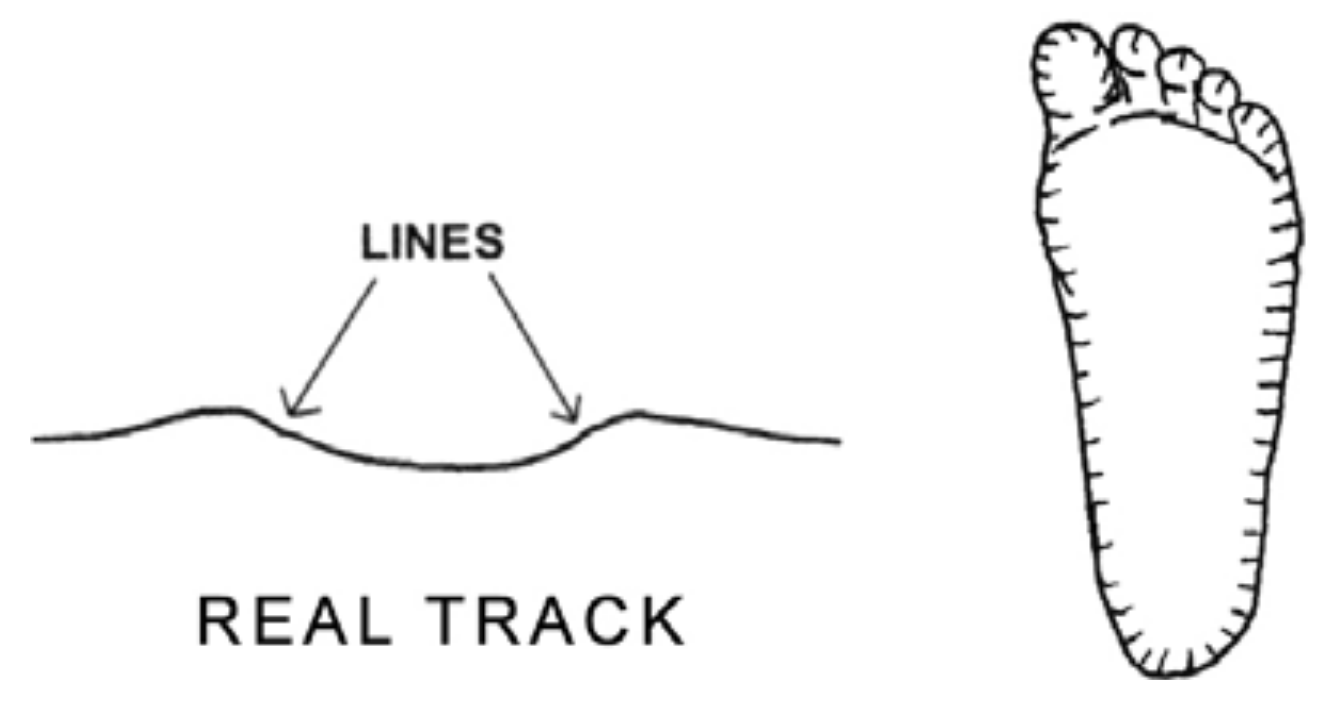 |
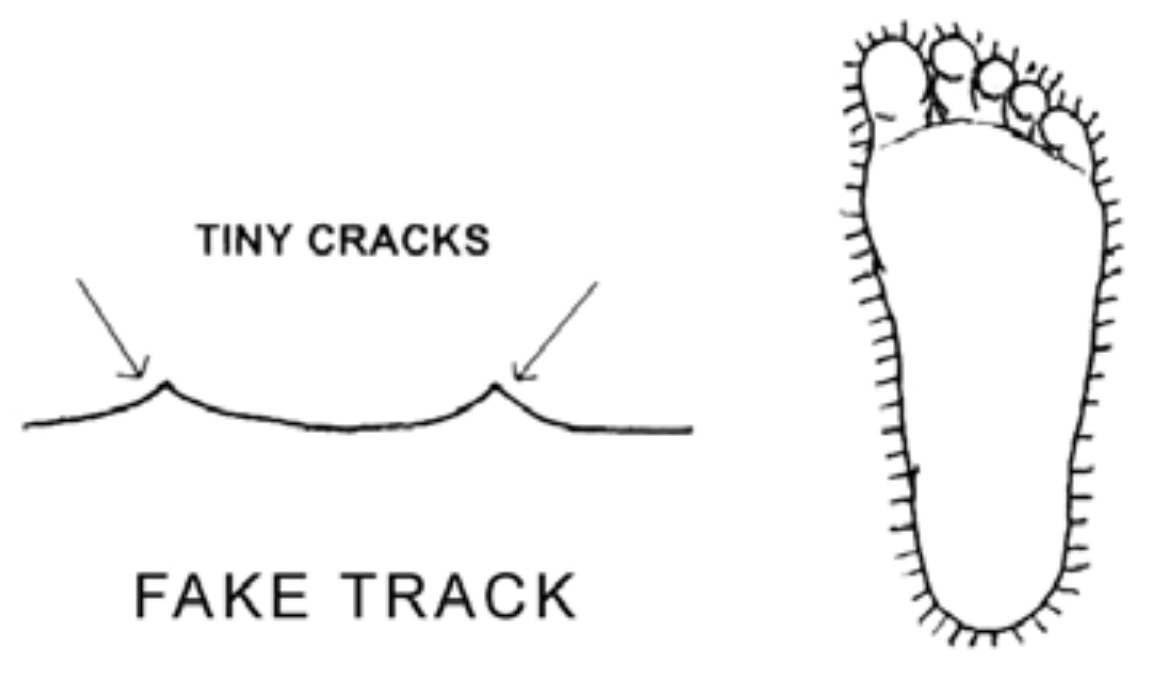 |
An entire branch of science is devoted to track analysis — ichnology. This science studies the tracks left in prehistoric rocks, and icnologists can tell an enormous amount about whatever made a particular track on a piece of stone.
Through nearly all of human history, tracking skills were a matter of life and death. In some isolated primitive areas today, it is still crucial to be able to “read” the tracks of all creatures in the local area to secure food for oneself, and to avoid becoming food for any of those creatures.
The fundamentals of tracking are simple. When a moving foot made of flesh and bone (a hoof would be different) is pressed into a medium with enough cohesion to leave an imprint (firm mud will work, loose mud won’t), a smooth and sequential progression is created by activating muscles, ligaments, tendons, and bones. This leaves an unmistakable “look” to a print. Tiny vertical cracks will form along its inside (see right). These cracks are compression lines.
Similarly, when any fake foot is applied to a sufficient medium, whether the fake is made of wood, plaster, plastic, or whatever, it cannot go down sequentially. It must be pressed down all at once, of a piece, leaving a distinctive “look.”
An easily noticed uplift — an impact ridge — is created along the track’s outside perimeter, and tiny parallel cracks will be found along it rather than the inside perimeter. A third-grader with a good magnifying glass would not be fooled by a fake, much less a qualified track expert.
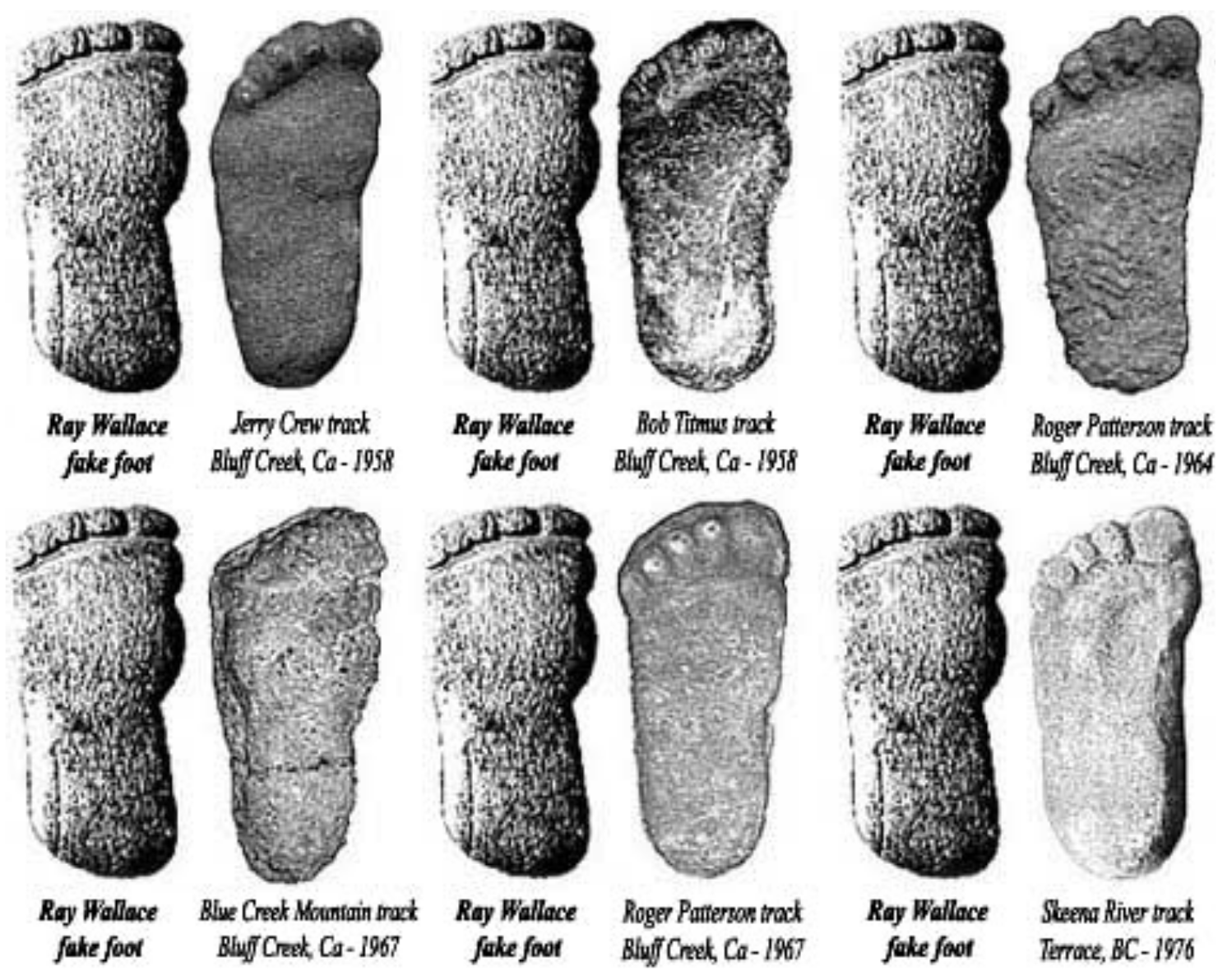 |
 |
Chris Murphy and all other researchers apply this kind of rigor to the analysis of hominoid tracks. Here is a comparison of six of the best hominoid tracks alongside the wooden tracks carved by a hoaxer named Ray Wallace.
Ray Wallace is on a par with Doug Bower and Dave Chorley, English barflies who claimed to have made 200 crop circles from 1978 to 1991.
When Wallace died in 2002, his family tried for their 15 minutes of fame and cash by claiming their patriarch had created all of the best-known bigfoot tracks, since the first ones discovered by a lumberman, Jerry Crew, at Bluff Creek, California, in 1958 (first comparison above).
In reality, thousands of hominoid tracks have been photographed and/or casted around the world during the past 60+ years. It started in 1951, when British mountaineer Eric Shipton photographed a line of unusual tracks (bottom right) across the Menlung Glacier in the Himalayas.
Because an Abominable Snowman/Yeti type made these, they are ape-like in structure, with large and opposed first toes, apparently for safe and easy purchase on narrow ledges of rock anywhere in the rugged Himalayan ranges.
 |
 |
Another fascinating aspect of hominoid track morphology has been discovered by Dr. Jeff Meldrum of Idaho State University, one of the very few credentialed academics with enough courage to speak openly about a subject his mainstream peers avoid whenever possible.
After analyzing dozens of hominoid footprints, Dr. Meldrum concluded that their feet are built very differently from human feet. This would make sense if they are bipedal primates that have existed on Earth for 20+ million years.
In dealing with so many tracks, Dr. Meldrum noticed that many of them leave a distinctive “break” across the middle. The photo (top right) is one of the best examples of this “break.”
His explanation for this kind of print is that the bigfoot foot must extensively flex in the mid-tarsal area in a way that human feet cannot.
Bottom right is an illustration that compares the foot, ankle, and heel redesign that would be required for a walking bigfoot to create such prints.
Providing excellent support to Dr. Meldrum’s observations is that gibbons have feet that work exactly the same way. This counters the long-held premise that humanity’s arched and rigid foot is an optimum design for walking upright.
 |
 |
Gibbons are arboreal, but when on the ground they can walk by holding up their long arms.
Careful studies of gibbons walking have shown they stride much like Dr. Meldrum’s projection. They have a “flexible” joint midway along the foot to support them when climbing or walking.
When walking, their toes hit the ground first, which stretches the toe tendons. At the stride’s midpoint, a gibbon raises its heel while leaving the forefoot on the ground, which creates an arch along the top of the foot rather than along the bottom, like Dr. Meldrum’s figure above.
Those stretched tendons store energy, and when the toe leaves the ground they recoil, releasing the stored energy and providing the necessary propulsion to push the foot and body up off of the ground to walk upright quite efficiently.
Since gibbon feet work that way, perhaps the Terraformers placed that design on Earth at the start of the Miocene as the model for all upright apes (hominoids) to follow. One thing is sure: all hominoid feet are very unlike human feet.
The late Dr. Grover Krantz of Washington State University also contributed heavily to the field of hominoid research, focusing as Dr. Meldrum does on the bigfoot’s foot and ankle structure.
Dr. Krantz found these (bottom right) proportional differences between a human foot and the bigfoot filmed by Roger Patterson and Bob Gimlin on October 20, 1967, at Bluff Creek in northern California.
 |
The Patterson-Gimlin film is one of the earliest, and remains one of the best, pieces of evidence in support of hominoid reality. This film has been scrutinized in as much depth and detail as the Zapruder film of the Kennedy assassination, and it is easily available to view on the internet.
The bigfoot female seen in the film is at left, above, in one of the film’s clearest frames. She is contrasted with a photo from a recreation of the filming scenario by a BBC show in 1998.
The BBC recreation clearly illustrated the sheer impossibility of producing a believable suit that could mimic the creature’s movements in 1998. If it could not be done in 1998, it seems safe to assume that also would have been true in 1967.
In the film, the bigfoot’s muscles ripple under the hair of her shoulder and thigh. No suit can mimic that. If it were glued to the skin, the skin would not ripple with natural movement. It is a perfect Catch 22 for would-be hoaxers, so the “man-in-a-suit” argument should be dismissed.
As the caption says, notice the difference in the length of the arms. Witnesses consistently say hominoids have arms distinctively longer than human arms, with hands that swing down near their knees, as opposed to human hands that swing at or near the middle or our thighs.
Those arms are a hallmark of the short-armed apes of the Miocene, the ones with arms the same length as their legs, the ones we are told moved on the ground as knuckle-draggers when it is obvious all they had to do is stand upright to move much more easily and comfortably.
An enormous amount of criticism has been put out by skeptics determined to confuse people about the Patterson-Gimlin film’s credibility. In answer to those critics, I can point anyone to an excellent body of work about the topic done by scientists who study the film itself, and to those like Daniel Perez and Chris Murphy, who study the details of the circumstances of the filming.
For this eBook, let’s simply try to disregard the harpings of critics and focus on the well-known facts that support its veracity. First, as noted above, a credible suit still cannot be made by Hollywood’s best model makers. The problems Roger Patterson and Bob Gimlin would have had to solve in 1967 remain unsolved today.
Second, the outsized dimensions of the female bigfoot — and especially the natural sway of her pendulous breasts — could never be matched by a human in a suit. Equally impossible would be matching a height of 6+ feet and weight of 300 to 500 pounds, based on the depth of the tracks she left walking across a hard-packed sandbar.
 |
At right are Bob Gimlin on the left and Roger Patterson on the right holding a cast of one of “Patty’s” 14" tracks. Patterson died in 1972, only five years after making the historic film.
I have met and talked to Bob Gimlin, one of the two men there when the film was made. After a long, detailed chat with him, I have no doubt at all that he has always told the unvarnished truth about what happened on that historic day.
The only people I’m aware of who don’t trust Bob Gimlin’s word are those who have never met him. He’s as straightforward as men come.
The next convincing proof of hominoid reality is called The Minnesota Iceman. Much more controversy surrounds this peculiar case than the Patterson-Gimlin film. Both were equally real, but Patty fell into the hands of people who were good at heart, while the Iceman did not.
A choking mass of criticism exists against the Iceman story because of the disrepute of the man who owned it, and because of numerous conflicting stories about how he came to own it.
When you clear away all the swill, the dregs I have come to accept are what follows. Much of it is undeniable fact, but other parts are based on my own opinions and judgments arrived at during extensive research and conversations.
***
In early 1967, a land-shark scoundrel named Frank Hansen allegedly went hunting for deer near his home in Rollingstone, Minnesota, near the border with Wisconsin. In the woods of the hunting area, he encountered a male hominoid.
If a hominoid in Minnesota surprises you, be aware that credible sightings of one or another of the three hominoid sizes (giant, man-sized, or pygmy) come from every U.S. state except Hawaii. Wherever deep forests, jungles, and swamps exist, hominoids can live in them.
No one is sure what Hansen saw. If it was a bigfoot, it was a juvenile. If an almas/kaptar type, it would have been an adult. Whatever it was, Frank Hansen said he shot it and killed it.
Dozens of reports are on record where hunters find themselves in position to shoot a hominoid, but invariably they say it strikes them as wrong in some fundamental way, and they don’t shoot. Frank Hansen was not that kind of individual.
He apparently shot the creature in its mid-back with a high-caliber deer bullet, which severed its spine and blew out a fist-sized circular exit wound in the middle of its chest. It fell down, paralyzed from between its shoulder blades.
Mortally wounded, the terrified creature lifted its left arm for protection against the assailant moving to its front to apply a coup de grâce.
Hansen fired a head shot that went through the left arm just above the wrist and entered the left eye socket, blowing out the back of its head and causing its right eyeball to explode and hang in a shred, like an empty sack, on the right cheek.
Hansen and at least one other person on the hunting trip (the likeliest candidate is his son) had driven out in a jeep or truck (story versions vary, as, ultimately, do nearly all stories about this huckster), so they loaded the body into it and then hauled it to Hansen’s home nearby.
 |
 |
As it turned out, Hansen had connections to carnival shows all over the U.S. (he was later called The Abominable Showman), so he knew how to ply a “freak show” attraction. He knew he had a winner with the “creature” he had killed, so he set to work to capitalize on it.
Before the body could stiffen, Hansen arranged for it to be put in a standard 7-foot floor freezer.
He filled it with water, froze it, then had it fitted with strip lights and a thick sheet of plate glass just above the top surface of the frozen water.
He then had the freezer installed in the back of a 16-wheel truck with the intention of taking it out onto the U.S. circuit of carnivals and state fairs to charge people to see his “creature.”
His first event was in May, at the start of the 1967 carny season. At the same time, he had a serious problem to deal with. “Bigfoot” had been well known since lumberman Jerry Crew found and casted its tracks in 1954, but by 1967 there was no consensus on what they might be.
Some believed they were an unknown primate, while others insisted they must be some kind of “wild” human. If the latter, then Frank Hansen could be charged with murder for killing it. To avoid that outcome, he developed a plan.
Howard Ball, a model maker at Disneyland in Los Angeles, claimed Hansen hired him early in 1967 to make a wax and rubber model of the creature he killed. That way Hansen could have a “double” to prove to authorities that a model was frozen in the ice. It was a very clever ruse.
From day one Hansen advertised his creature as a “mystery” that even he didn’t have an answer for. He just put it on display for people to pay a quarter to view it … a typical carny spiel. (The price rose steadily as years passed. Nine years later, 1976, when I saw it, it was $1 per view.)

Frank Hansen with the fake Iceman
created by Howard Ball |
With his model available at a moment’s notice, Hansen could always seem agreeable when any authorities wanted to examine his creature. And the FBI supposedly did “lower” itself to contact him about it, but that contact is now in dispute.
Supposedly, Hansen showed them the copy in its ice tomb, they X-rayed it, realized it had no bones, and reported it as a fraud. Whether this story is true or not, other U.S. authorities soon lost interest in it. As Hansen had hoped, he was free to continue showing the far more realistic-looking real creature, safe from legal hassles.
Hansen knew that X-rays were the key, so he never allowed the real one to be X-rayed. He kept that aura of mystery about it as much as he could, making up various stories to avoid any fingers pointing to him as a possible murderer.
Then, after the 1967 carny season ended, on October 20, Roger Patterson and Bob Gimlin shot their film at Bluff Creek and stirred the pot of interest in Frank Hansen and his creature.
 |
 |
The next carny season, the summer of 1968, Hansen took it to events in Minnesota, Illinois, Wisconsin, Michigan, Oklahoma, and Texas. In that time, it came to the attention of two world-famous zoologists, Dr. Ivan T. Sanderson, and his Belgian friend, Dr. Bernard Heuvelmans.
(At right are Heuvelmans left, Sanderson right.)
In mid-December of 1968, Drs. Sanderson and Heuvelmans were able to view the exhibit in great detail at Hansen’s home in Rollingstone.
Both men wrote official reports, and Sanderson popularized it in a story in Argosy Magazine, May, 1969. (I have a rare copy of the issue, so illustrations in it will be presented here soon.)
Sanderson’s publicity led to the worst crisis Hansen faced with the Iceman. Upon taking it into Canada in July, 1969, Canadian Customs officials impounded it and demanded that it be X-rayed to prove it wasn’t a dead “wild” man.
Since this was the real one Hansen had killed, he had no choice but to refuse. Customs then initiated legal procedures to force Hansen to submit to the X-ray process. Being the shrewd man he was, Hansen had an ace up his sleeve.
 |
He had donated generously to his Senator in Congress, Walter “Fritz” Mondale, who in 1976 became Jimmy Carter’s Vice-President. He called Mondale and explained his problem.
Mondale made a call to Canada’s Customs, and Hansen was released, never to risk that again. He felt safe in the U.S. and stayed in the U.S.
 |
The model of the creature that Frank Hansen had made was good for 1967, but it would not fool anyone who had seen the real one. Many of the finer details were wrong or missing, and the “clouding” of the ice was in a different pattern.
It was, however, more than good enough to fool authorities, who went into their examinations of it with the typically strong bias that it must be a fraud. In the end, Hansen gave them what they expected to find, and they went away satisfied.
This was not the case with Dr. Ivan Sanderson and Dr. Bernard Heuvelmans. When they came to Hansen’s home in Rollingstone, Minnesota, he was seeking publicity and knew Sanderson could give it to him. Sanderson was often on national TV at that time, a bona fide “star.”
Frank Hansen knew he needed the right kind of publicity, so he demanded that they downplay, and not show, the incriminating bullet wounds. They agreed to his conditions, but grudgingly.
In photos (at right) the circular chest wound was obvious. But in drawings the scientists did for early reports, they left out the chest wound while depicting the wounded left arm as it was.
 |
Hansen gave Sanderson and Heuvelmans full access to the Iceman’s frozen corpse, allowing them to study it for hours under strong lights so they could carefully evaluate it and form their opinions backed by long years of experience.
Below is an expanded version of the sketch, along with detailed measurements Sanderson and Heuvelmans made to show how the body fit into its 7-foot floor-freezer “coffin.”
As the diagram makes clear, it was larger than humans in every dimension. Bent knees make accurate judgment difficult, but 6' 3" is likely. And those bent knees were an oddity. They had room to lie flat but did not, as if they couldn’t simply lie flat. This will be significant later.
The text Sanderson wrote to go with this drawing is:
“The creature’s characteristics include short neck, arms reaching to knees, disproportionate hands and feet, and [an] extremely husky body.”
These features agree with what is known of the classic Neanderthalers. Some scientists, mostly Russian and Mongolian, have held for years that scattered populations of these prehistoric men still survive in remote areas.
Dr. Bernard Heuvelmans wrote:
“The specimen is an adult human-like male, six feet [or more] tall, differing from all types of modern man by these striking characteristics [below]:”
- Extreme hairiness (covering the body).
- An apparent shortness of the neck.
- A barrel-shaped torso, more rounded than (in) modern man.
- Extremely long arms, which must reach to the knees when hanging.
- Disproportionate hands and feet. Hands are 11" long and more than 7" wide.
- Peculiar relative proportions of both fingers and toes. The thumb is longer than [a] modern man’s and the toes are all nearly the same size.
Anybody doubting that this historic relic was ever evaluated by serious professionals of the highest caliber needs only to read their articles.
***
 |
 |
 |
The water covering the body was not purified, so it froze haphazardly. Half was crystal clear, like looking through 6 to 18 inches of air.
Swirled through mostly the left side of it were the milky clouds that form as tap water freezes. (Think of ice cubes from a refrigerator. Some parts will be sparkling clear, while others will be milky.) These were translucent or opaque.
  |
Despite the obscured parts of the ice (depicted left), the clear parts were over the most important areas. The face was clear. The shot wrist was clear. The chest wound was clear.
But, in a seeming stroke of modesty, most of the pubic area was veiled by the milky clouds.
In the original drawing, the wound in the chest was included. In the refined image for Argosy Magazine (left, on right), the obscuring ice was left out to make a more viewable image. Also, the chest wound was erased, and the nose, hands, and feet were reduced to appear more human.
Because of the way the body rested in the ice, certain parts of it were much clearer than other parts. Heavily calloused kneecaps (indicating it spent a lot of time kneeling), and the left palm and right forefoot were especially clear.
The right foot was measured as 10" across, the size of an average dinner plate. It was simply a very large being, by any scale of dimension.
I was fortunate enough to see the real Iceman in 1976, and I was able to go through the viewing line three separate times. I was thirty years old then, and I had exceptional eyesight, what my father, an optometrist, called “astronaut eyes” because they equaled the astronaut standards.
By then I was also familiar with the subject of hominoids, having read whatever I could about them whenever I could find information. (This was well before the internet, so news about this subject was not easy to come by routinely.)
The first thing I noticed was how clean it was. Not a twig or a sprig of grass could be seen in its hair, and all of what had to be an enormous amount of wound blood had been sprayed clean by hosing it from the hair covering the body.
Speaking of its hair, I was deeply impressed by the stunning precision of the myriad follicles I could see. Because the body rested in the water until it froze, the water lifted them off the skin, making viewing perfect in the clear-ice areas.
I would guess a million hairs were on the body, each 3" or so. Every hair was perfectly round and noticeably thicker than typical human hair, the size of thin leads put in mechanical pencils.
 |
|
[Below are magnified agouti hairs from a cat.]
 |
Each hair was neatly tucked into a tiny dimple in the skin, and spread across the skin with odd consistency, like birds spaced along wires. Fake hairs “punched” into latex skin always exhibit a mark made by the tool that punches in the hairs, and consistency of spacing is flatly impossible.
[Discussions of the problems encountered using this technique come from the expert modelers at Madame Tussaud’s famous wax museums.]
Each hair was symmetrically banded from tip to bottom, dark alternating with light, with larger bands at the base and thinner bands at the tips.
These are distinctive traits in the hairs of certain animals, called the agouti pattern, which some monkeys have but not apes or humans. It would be excruciatingly difficult to fabricate a million such hairs with such precision, now or in 1967.
Each hair was straight or at most gently curved. Because so many of them had floated up off the skin, the skin was seen as pasty white, not dark.
Diametrical to its growth in humans, no axillary hair was visible in the armpits or in the crotch. To create a fake with such perfection at every point where hair entered the skin, was — and to my mind remains — utterly impossible.
 |
For as convincing as the hairs were, what sold me on the Minnesota Iceman’s veracity was the wounds caused by its killing. Each was distinct and clear, making them easy to see and judge.
[If you are creating a fraud of this nature, why mutilate it? Or, for that matter, if the Patterson-Gimlim film was a person in an ape suit, why bother to create realistic pendulous breasts?]
The most obvious wound was the bullet’s exit hole in the chest. It would have neatly held a baseball. Its edges were frayed and colored pink-red due to ruptured capillaries and veins.
Rising up from inside it were several twisting tendrils of pinkish plasma, each a few inches long and the width of a soda straw, and all of which floated upward before the water froze.
When all tendrils reached the water’s surface, they spread out into different-sized circles as the water froze. The largest was the size of a saucer, the smallest like a quarter or a nickel. Every other wound produced the same kind of straw-sized, pink-tinged, ribbon-like tendrils.
The collarbones arced up to an unusual degree because the rear of the skull was blown away, causing the head to rest lower than it normally would have. This left the clavicles bowed and stretched up-and-over at an abnormal angle.
The mouth was slightly open, and parts of teeth were visible. They were covered with diluted blood, making them a darker pink than the one tendril weaving upward from each corner. The nose was quite wide and pugged like a gorilla’s, but definitely fleshier than a gorilla nose. Each round nostril sent a tendril rising to the surface.
The left eye took the bullet and was simply a dark hole, like the chest wound, with its own two or three tendrils coming up. The right eye, too, was an empty hole with its own tendrils rising up. However, the remains of its empty eyeball rested on the lower outside part of the eye, like a large teardrop added on for effect.
As the illustrations show, the left arm was thrown up and rested above the head, so the terrible wound to the forearm above the wrist was very clear beneath its few tendrils. White chips of shattered bone were visible in a jagged jumble within and just outside the gaping hole.
The chest was barrel-shaped, much more than a human, and a bit flared out at the lower ribcage. The hands and the feet were extraordinary, too, much larger than humans, and uniquely shaped, with long thin thumbs, and toes that were more or less the same size rather than diminishing.
To me, to my mind and eyes, it gave a sense of overwhelming physical power. I played football through college as an undersized running back, so I have a good sense of what physical power is, and this creature, even dead, bristled with it.
So, what became of it? Nobody knows. Some people say it was decaying in the ice, and that it reached a point where Hansen could no longer take it out and show it. But when I viewed it in 1976, it looked fine and did not smell of death.
If Frank Hansen had wanted a better footnote in history, he could have turned over the remains to some favored university, but he most likely buried or burned it. Since his death in 2005, most researchers assume it is lost to history.
Speaking of researchers, several have tried hard to convince me that the Minnesota Iceman can not be taken seriously because of the many wild stories Frank Hansen told. But, stories or not, I know damn well what I saw that day in 1976.
An old saying in baseball is that a no-hitter is a no-hitter, even against Lighthouse for the Blind. Likewise, a dead hominoid in a block of ice is a dead hominoid in a block of ice, no matter who thinks, feels, or believes it was not.
I’m positive that anyone who examined the real Iceman will not be among those who dismiss it as irrelevant. If you ever saw it, then you know.
 |
Zana was a female almas (almasty in Russian dialects) type of hominoid. She lived and died within memory of many people still alive in the Russian village of Tkhina in the early 1960s.
At that time, Dr. Boris Porshnev was Russia’s equivalent of Ivan Sanderson, a highly qualified academic who also investigated hominoids as a matter of personal interest. Porshnev spoke to dozens of people who knew Zana well when they were children in the village of Tkhina.
Tkhina is in Abkhazia, a Kazakhstan province near the eastern coast of the Black Sea, along a southwestern flank of the Caucasus Mountains. It is on the Mokva River, and is 78 kilometers from Sukhumi, the region’s capital city.
Locals are very long-lived, with many reaching well over 100. Porshnev found ten people who attended Zana’s funeral around 1890, but they could not agree on the year she died, or on her burial spot in Tkhina’s cemetery. It’s a Muslim area of Kazakhstan, so graves were not marked.
No one knew how she was captured, or when, or how old she was at the time, but they agreed she lived in Tkhina for about 40 years. Hunters would capture male almas to train as slaves to do hard physical labor because they were so incredibly much stronger than humans.
Captured female almas also did hard labor, but in addition were used as sex slaves for human males, and as wet nurses. Such captures and enslavements were rare, but they were well- recorded events in the 1600, 1700, and 1800s.
Upon capture, Zana was wild with rage, forcing constant shackling and frequent gagging with felt to muffle her loud screams. She was passed from one owner to another, until she was given to Edgi Genaba, a man of high status in Tkhina.
Genaba carted her home, bound and gagged, and put her in a stout enclosure near his house. She dug a hole in the ground and lived in it for three years, fed through cracks in the enclosure.
Eventually her anger began to subside, and she became tamed enough to wander around on her own. By then she was like a “pet” that always came back to where she was fed and where she slept, which was a hole she dug under an eave overhanging the roof of Edgi Genaba’s house.
Today, the top experts about Zana are Russian hominologists (the term they prefer) Dmitri Bayanov and Igor Burtsev. Both men were with Porshnev in Tkhina. When Porshnev died in 1972, they assumed leading roles in this vital area of research. The following is derived from their years of effort to document Zana’s story.
***
Zana was described as tall and thick-set, with heavily muscled arms and legs. Her body was covered with long, reddish-black hair. She had enormous bosoms and buttocks, and her hands had fingers that were longer and thicker than in humans. Her feet were also large, and she could splay her toes widely, especially the big toe.
Her face was broad and clear of hair, as were her palms and the soles of her feet. The skin on them was gray, and her face skin was dark gray.
[Relative to the Minnesota Iceman, with a full portion of blood still in his body, the white skin in the freezer may well have been a shade more like the darker tones of Zana and the bigfoot in the Patterson-Gimlin film, which is the norm for sighting reports of hominoids worldwide.]
She had high cheekbones; a wide, flat nose with nostrils turned up-and-out (pug); a muzzle-like (prognathous) mouth that was wide, with large white teeth; a low forehead above a heavy brow ridge, and dark eyes with a “reddish” tinge.
The reddish tinge indicates a much larger retina area, filled with many more capillaries than are found in humans. Animals with such eyes can usually see well at night, and Zana often went for long evening rambles across hill and dale.
According to the locals, Zana’s most arresting feature was the normal expression of her face, which was distinctly animal rather than human. Sometimes she would emit a loud, spontaneous laugh, baring her big white teeth, which were strong enough to crack the hardest walnuts.
Her head hair was thick, tousled, and stood up on her head like a top hat. It was long enough to hang like a mane down her back. She always had a pungent body odor, despite often bathing or swimming in the nearby Mokva River.
She lived in Tkhina for about 40 years, showing no physical changes: no gray hair and no falling teeth. She was vigorously active throughout her life, which kept her strong and fit. Her athletic abilities were, and to the end remained, superb.
She could outrun a horse at full gallop, which is normally about 40 mph. Humans reach 22 mph.
She could swim the wild Mokva River in a full tumult of flooding during the spring thaw.
She climbed trees to get fruit and would gorge herself with grapes when they were in season. She ate whatever was offered, including meat and hominy, with bare hands and great gusto.
She loved wine and often overindulged, after which, like most of us, she’d sleep for hours.
She preferred to walk naked, even in winter, always exhibiting an imperviousness to cold. She would tear into shreds any dress she was given. However, in later life she tolerated a loincloth, for whatever modesty it provided.
She couldn’t speak, apart from uttering various murmurs, yelps, howls, and growls. In her 40 years in Tkhina, she never learned to speak a word of the local language. However, she was clever enough to learn that language and could understand whatever she was ordered to do.
She had acute hearing, and did what she was told. Her primary chores were gathering and carrying the wood for village fires, an onerous chore for village women that Zana relieved.
The same was true for hauling water from the nearby Mokva River, another strenuous chore that Zana had to perform daily, but it was easy because of her extraordinary physical strength.
Her great strength was also used doing men’s work, especially carrying heavy sacks of grain or meal (reputed to weigh up to 80 kilograms, or 180 pounds) to and from the nearby mill.
She was entirely a slave, and lived as such.
Zana never liked dogs and they did not like her. (This is consistent with all types of hominoids, perhaps reminding them of wolves). She would toss sticks and stones to keep them away.
 |
Children would often tease her in various ways, but she never hurt any child who bothered her.
She could not tolerate being in heated rooms, preferring to stay outside in the coldest weather. As mentioned, she often cavorted in the Mokva River, immune to its bone-chilling temperature. She clearly had a very high tolerance for cold.
She often played — sometimes obsessively — with rocks, grinding them together, or flaking and smashing them into chips and cores. She seemed driven to create edges and/or points on stones, which strongly resembled what are now called the Mousterian “tools” of Neanderthals.
However, she never used them for anything more than projectiles for throwing at dogs.
 |
The most astounding part of Zana’s story is that on eight occasions she was made pregnant by men in Tkhina, giving birth with no difficulty or assistance. Then, following primal instincts or her species’ protocol, she’d carry her hybrid newborn to the river to clean it, and the shock of being dunked in ice-cold water would kill it.
That happened with her first four births, all of which looked amazingly human, as if her role in their conception was extremely minimal. So after her fifth birth, the village women took the infant from her to raise as human because every new pair of hands in any poor rural village like Tkhina could grow up to be a valuable asset.
Zana’s final four children — two daughters and two sons — lived because they were taken from her before she could wash and inadvertently kill them. They all grew up to be much more human than whatever she was. They looked and acted like normal people, with full powers of human speech, normal intelligence, and sound reason.
Naturally, certain of their mother’s traits were in them. All four were more robust that usual, much stronger, had darker skin and rougher facial outlines, but all were basically human.
Zana’s eldest son, Dzhanda, and her eldest daughter, Kodzhanar, were raised to adulthood and then left Tkhina to live in other villages in Abkhazia, where they supposedly raised typical families. Her second daughter, Gamasa, lived in Tkhina and died in 1930, and her youngest son, Khwit, did the same, dying in 1954 at about 70.
Zana was buried in the Genaba’s family plot, as later was Edgi Genaba and his wife, who raised Gamasa and Khwit as her own. Gamasa was buried near that plot, and then Khwit near her.
 |
Boris Porshnev exhumed a skull in 1965 that is believed to be Gamasa’s, while in 1978, Igor Burtsev did exhume Khwit’s skull (see below).
 |
Locals recall Gamasa and Khwit as robust and powerfully built, with dark skin and vaguely Negroid features, but without overabundant body hair. Human traits were strongly dominant in both in every meaningful aspect, which is not how species hybridization usually works. (This will be addressed in much more detail later.)
Khwit was remembered best by everyone the Russian investigators interviewed. He had dark skin and thick lips. His head, which was large for a human, was small in relation to his robust body. Though not Negroid, his head hair was stiffer than usual, and stood up straighter.
He was extremely strong, and tended toward a contentious personality. He often got in fights, and lost his right hand in a fight, but could still work with his left hand, and even climb trees! He had a high-pitched voice and sang well.
***
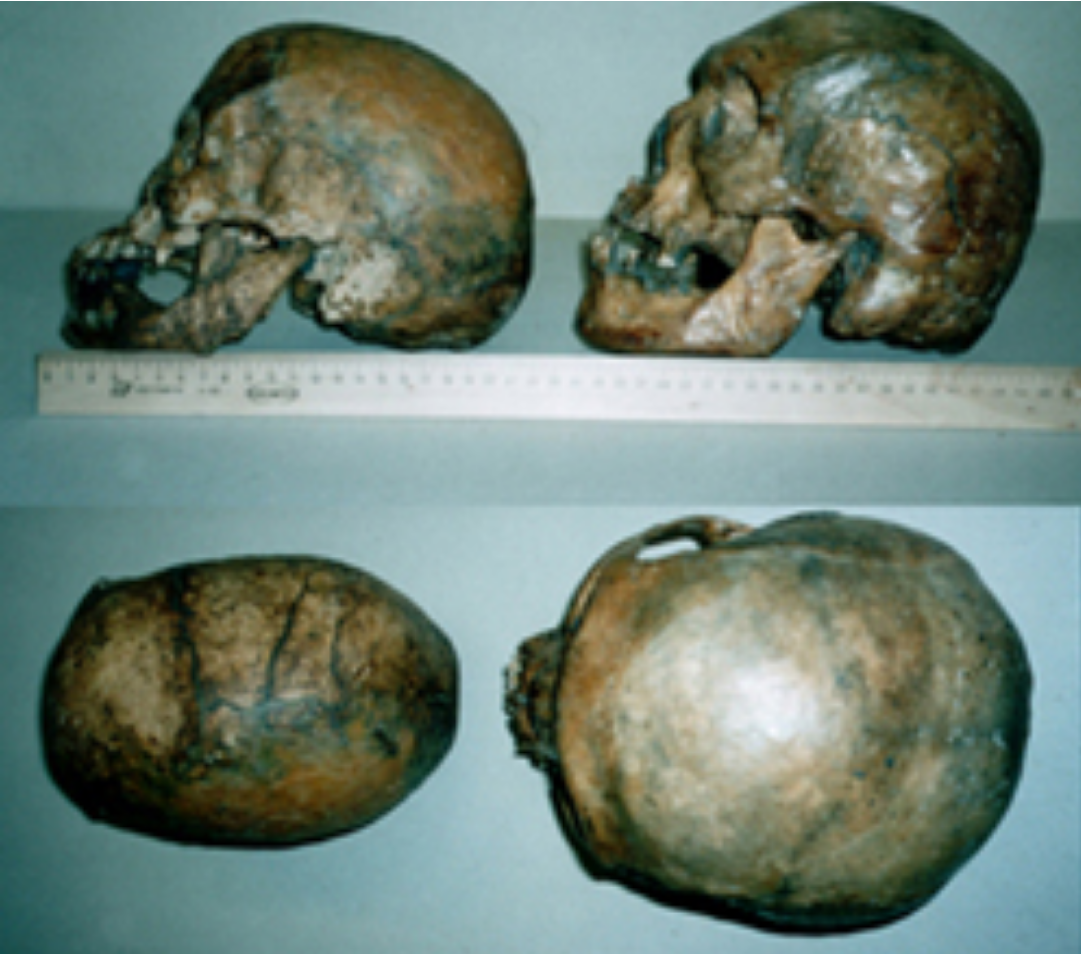 |
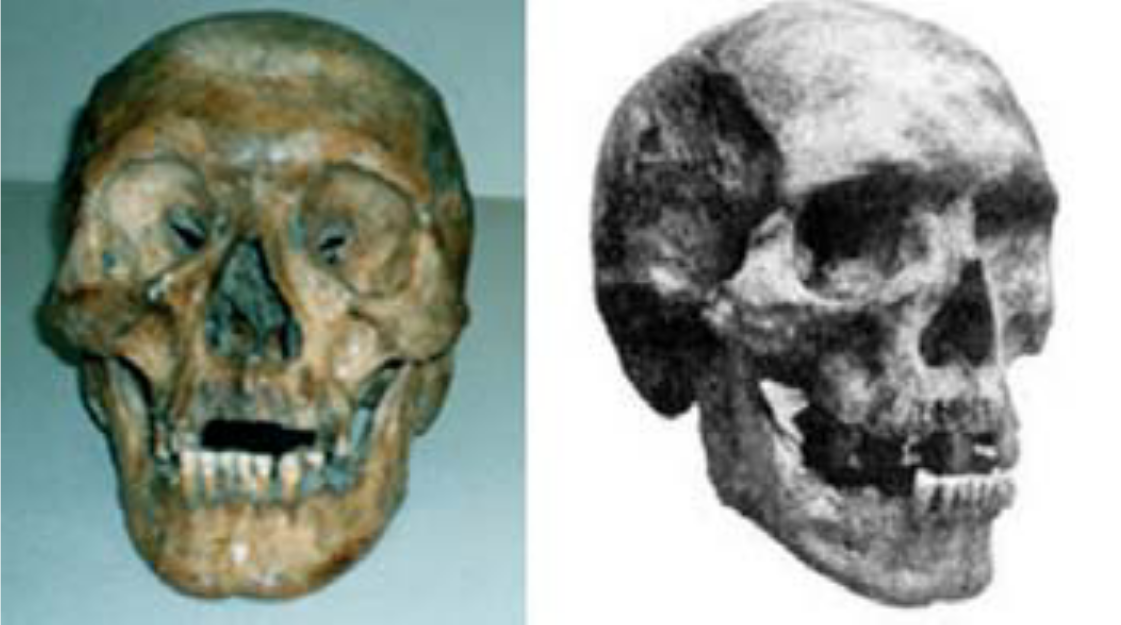 |
 |
Khwit’s skull shows very strong features for a human. It is on the right, compared to a human female’s skull (on the lrft) found buried near him.
Some experts, like the late Dr. Grover Krantz, argued that Khwit’s skull remained within the range of standard human variation, but others make the case that he was significantly past it.
 |
 |
 |
Igor Burtsev also sought out and located three of Khwit’s children, and some of their children. Khwit’s oldest daughter, Tanya, was 53 years old in 1971. The photo at left is one that was current then, and one when she was younger.
(Middle left) Khwit’s son, Shaliko, and daughter, Raya.
Shaliko’s daughters, Rita and Zoya — Zana’s great-grandchildren — taken in 1976 (bottom left).
(Bottom right) Khwit’s wife, Maria, left, his daughter Raya, right, and his granddaughter, Manana, center.
In studying the Zana case, let’s make clear that her descendants exist, and those that descend through her daughters carry her mitochondrial DNA, which is passed down through females.
Why haven’t mainstream geneticists tested any of Zana’s female-line descendants to see if their mtDNA is significantly different from a normal human? Maybe they already know the answer.
It is hard to estimate how many times I’ve been asked a question along these lines: If hominoids are real, where are they? Why don’t we already know about them? It’s not like we don’t know everything else about our planet, is it? Look at Google Earth! Nothing can hide from that!
I know they mean well, and so do you if you’re thinking, “Yeah! Where are they?” The truth is, they are everywhere, on every continent except Antarctica, and able to live and flourish in any deep forest, jungle, or swamp in the world.
If you live in any U.S. state other than Hawaii, one or another of three hominoid types (not the yetis) have been in, and could be in, your state.
If you live anywhere in the world with suitable habitat for them, they might well be there. I do realize this is a mind-bender, but there it is.
Here is another: How much of Earth’s surface do you believe remains unsurveyed? Not by Google Earth’s satellites, which are only good for above-tree-level viewing. I mean on the ground, foot-surveying, where every hill and dale is registered and recorded? How much?
I will save the details of this answer for a later eBook strictly about hominoids, but here is the bottom line: A conservative estimate is that about 30% to 40% of the Earth’s land surface — which includes the polar ice caps and all of the deserts — has been foot-surveyed at least once.
That is 30% to 40% of 58 million square miles (M/sq/mi), or 17 to 23 M/sq/mi. Much of that surveying was inadequate once-overs carried out by early explorers moving through rugged areas for the first — and often the last — time.
In the U.S., 380 million acres of western federal land has never been foot surveyed. That is 17% of the entire U.S.! Another 50 million acres — 2.2% of the U.S. — were last surveyed, poorly, a century ago. Thus, we know next-to-nothing about 20% of the western states in the U.S.!
Since the advent of flight, the vast majority of land surveying has been done from the air — first by planes and then by satellites — because dense forests and swamps are so difficult to get through on foot carrying surveying equipment.
Not only is 30% to 40% of Earth’s land all that has been surveyed, only about 20% of that — 3.5 to 4.5 M/sq/mi — is considered well-surveyed, and that’s due almost entirely to oil exploration.
Thus, the 60% to 70% of land on Earth that has not been surveyed is basically terra incognita!
Don’t misunderstand and assume I’m saying hominoids run around on 60% to 70% of Earth. That isn’t what I meant. Like all species on the planet, hominoids have their ecological niches where they are comfortable living, and humans have our niches where we’re comfortable. The key is to understand those niches are different.
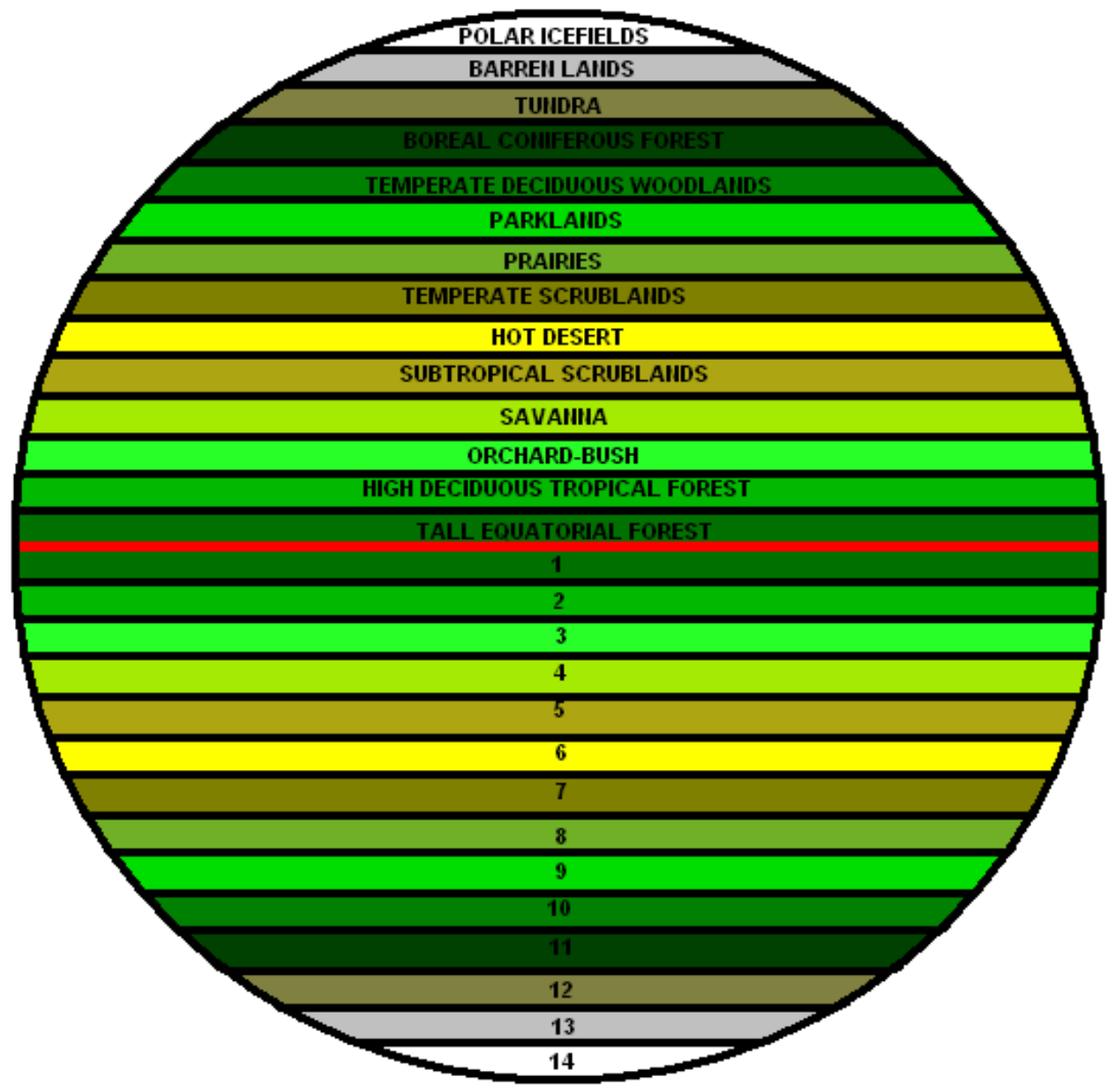 |
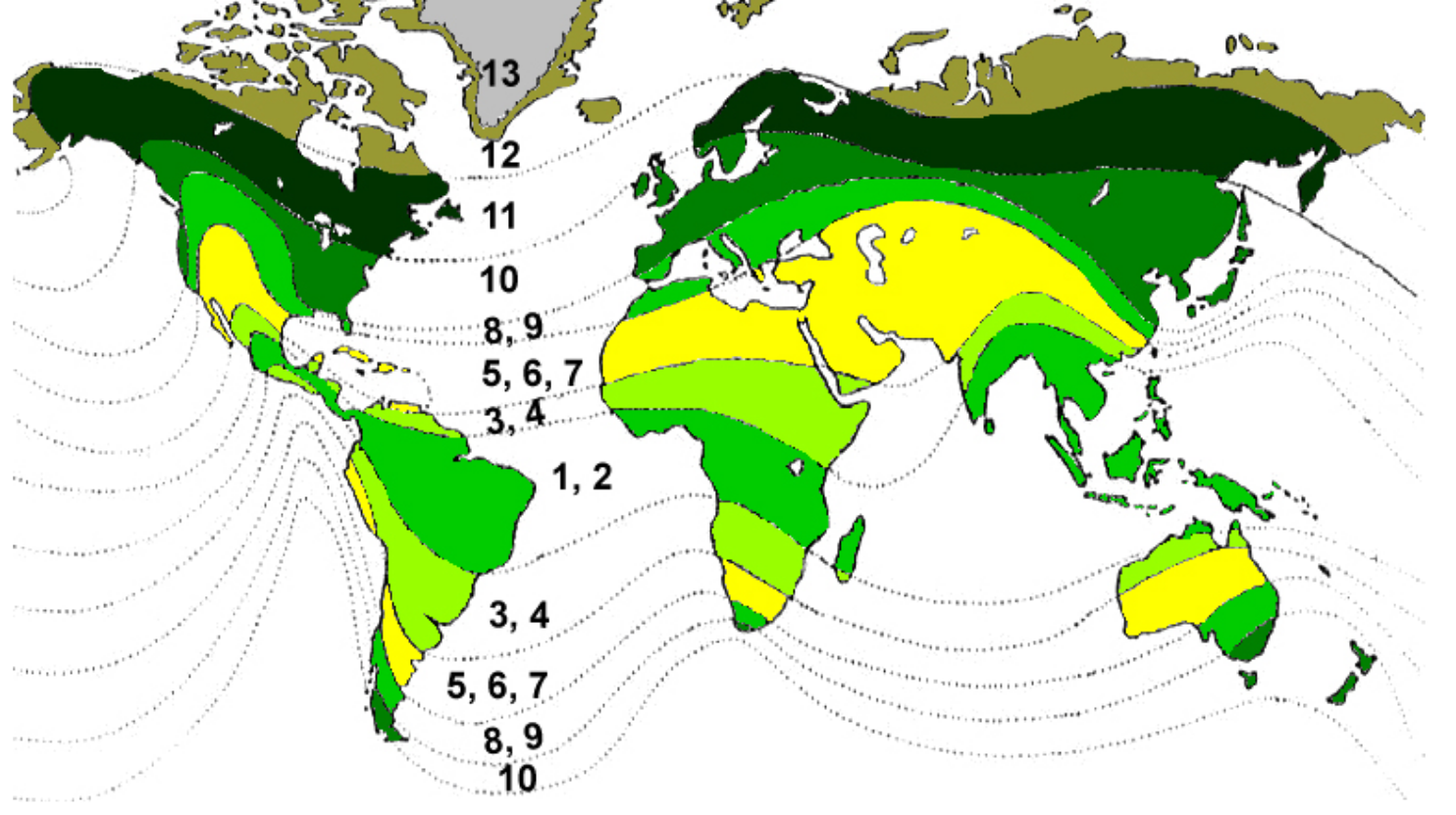 |
If Earth were a perfect sphere with no axial tilt and no mountains to disrupt weather patterns (below), you could start walking at the equator, heading north or south, and pass through the same 14 ecological niches in either direction.
You would pass through a new niche every 900 miles (1/14th of the hemisphere circumference, 12,600 miles). Obviously, Earth is not a smooth ball with ideal weather patterns for vegetation.
Real-world environments are shaped by ocean currents that drive winds, which then encounter mountains that distribute or hinder rain, which in their turn distribute or hinder all vegetation. Nonetheless, a surprising degree of regularity can be found across the 14 ecological niches.
The ecological niches thus created sweep from either side of the equator, to north and south, in the same predictable patterns. Few people are aware of how consistent these patterns are.
Starting at the equator of real-world Earth and moving north or south, the bands we find are:
- Tall Equatorial Forest
- High Deciduous Tropical Forest
- Orchard-Bush
- Savanna
- Subtropical Scrublands
- Hot Desert
- Temperate Scrublands
- Prairies
- Parklands
- Temperate Deciduous Woodlands
- Boreal Coniferous Forest
- Tundra
- Barren Lands
- Polar Ice Fields
The prime ecological niches for humans are the orchard-bush and savanna, and the prairies and parklands. We live in others as well, but never in large numbers. We cluster in four (4, 5, 8, 9).
By far the sparsest human populations are in the deserts and scrublands, the tundra, barren lands, and polar ice fields. The hominoids own all of the rest of it, the forests and the woodlands.
I know, we do go in those areas, we work there, we cut many forests down. But we don’t live in those areas, we don’t cluster there. Our urban centers — all combined — cover less than 2% of Earth’s land surface — only 1.2 M/sq/mi. If that statistic doesn’t get your attention, it should.
***
Earth’s total land surface is 58 million square miles. Of those, 33% (19 M/sq/mi) are the icy wastelands and the waterless tracts of deserts unfit for human habitation. (Desert dwellers have little choice but to live around oases or near sources of fresh water, and they usually move in straight lines from oasis to oasis.)
Apart from the ice caps, tundras, and deserts, another 30% of Earth’s land (17.5 M/sq/mi) is covered by temperate forests. It includes thick montane growths of hard and soft woods, dense jungle canopies, and swampy bottomlands.
While huge tracts of forests (the Amazon basin is an enormous one) are very thinly populated, as are deserts like the Sahara, Gobi, etc., in real terms such vast areas are virtually uninhabited.
Apart from ice caps, tundras, deserts, and dense forests what remains — 37% (21.5 M/sq/mi) — is arable land on which humans live comfortably. Let me repeat that: Only 37% of Earth’s total land surface is easily and readily available for typical human habitation. Surprising, isn’t it?
What it means is that we humans are quite far from being the masters of all we survey. We do not even survey all we claim to be masters of! Our vaunted “mastery” extends to only a shade more than one-third of Earth’s total landmass.
Another third is too frozen or parched to be of much use to anybody. And the last third clearly and without question belongs to the hominoids.
Hominoids have plenty of room to live their lives — 50, 60, 70 years, whatever they are — without ever seeing a human. If you doubt this, check with people who live on the edges of the overlaps between our niches and their niches.
Those places consistently produce the rare encounters with hominoids. People who live there know they are real because they hear, smell, or see them—and sometimes all three.
The humbling truth is that human ecological niches do have restrictions, as do the niches of all the other creatures that live and die with us. This is a law of Nature hominoids easily obey.
Another question I’m often asked is this: “If Hominoids are real, why don’t we see them more often? Why aren’t we finding their dead bodies on a regular basis? Hunters are out in forests all the time! It doesn’t make sense.”
They’re right, it doesn’t make sense, but only because they don’t know how animals actually die in the wild. Ask anyone with a long career in forest environments: “Have you ever come upon the corpse of a non-prey animal that laid down and died in an open space where it was easy for you to see it or its remains?”
The answer will be a resounding “No!” Seeing the partially eaten remains of a prey animal is common in forests, but seeing one of any non-prey animal is so rare, it almost never happens.
 |
With the exception of the agogwe-sedapa types that live in jungles and would be prey for large jungle cats, the other three hominoid types are not likely to die from attacks by other animals. Their juveniles, yes, of course, but not adults.
The three larger hominoids have overwhelming physical strength (remember, pound for pound, all primates — including monkeys — are 5 to 10 times stronger than humans). They also have an ample dose of intelligence, and are the absolute masters of their various rugged environments.
The only way larger types might leave openly visible remains would be if they were shot (a few such cases are recorded), or hit by vehicles (a few of those, too), or otherwise the victims of untimely accidental death (a fall off a cliff).
Like all other non-prey animals, the larger hominoids will reach old age, sicken, and die. They will know when their time has come and will seek out a secure, well-hidden shelter to tuck into and spend their last days and hours.
 |
Living as they all do, in wooded terrain on flat land or mountains, they can easily find pockets of rocks or fallen trees to serve as a makeshift “coffin” while they wait out their natural end.
When it comes — soon, or after an agonizing wait — Nature swiftly moves in to clean up the remains. Scavengers will get to fleshy parts and strip the body to its bones. Insects and worms will clean out what remains of marrow, etc.
The bone will be attacked by molds and fungus that utilize certain parts of it; and what remains will be lying on the acidic soils of all forests.
In weeks the soil acids will dissolve every scrap down to nothing, leaving a “clean plate,” so to speak, giving no hint anything ever died there.
A key point to keep in mind is that the forested areas supporting hominoids are the very same environments that have produced only a few fossilized teeth of any chimpanzees, and most bones alleged to come from gorillas, monkeys, baboons, and orangutans are subject to dispute.
The processes that keep forest inhabitants out of the fossil record are what act to prevent easy access to hominoid remains. So just because we don’t find hominoid corpses lying around in the woods, it does not necessarily mean they don’t live — and finally die — there in large numbers.
 |
 |
The Panda’s story is a perfect comparison with hominoids because they live in the same kind of habitat (montane forest) and terrain (mountains) as bigfoot/sasquatch and the almas/kaptar types, but their mountains are covered with bamboo.
Written references to hominoids go back for hundreds of years, and such mentions are found in nearly every country. But the mainstream of each century has dismissed them as useless prattle. The same was once true for Pandas.
Ancient Chinese manuscripts often mentioned a creature known to them as a Bei-Shung, which meant white-bear. They were usually described precisely as they turned out to be: black-and-white bears living in the rugged mountains of Sichuan, eating nothing but bamboo shoots.
Every Western authority “knew” bears were omnivores, so the bamboo shoots were laughed off. Also, they “knew” bears could be black or brown or white, but surely not the black-and-white coloring described by Chinese natives.
Most importantly, no Western authority had ever braved the rigors of traveling to China to study the matter, so they felt amply qualified to pronounce the Panda an amusing local legend.
Finally, after 2,000 years of unaccepted reports, in 1869 a French missionary/naturalist named Father Armand David made his way to Sichuan Province. Like most naturalists, he knew of the legendary Bei-Shung, and he believed “official” assurances that duotoned, bamboo-eating bears were a highly embellished Chinese fable.
Ultimately, Father David saw the full skin of one hung on a wall in a village elder’s home! Finding out where it came from, he hurried to the Bei-Shung’s reported habitat, a bamboo forest high in a forbidding mountain range.
Upon arrival, he tried to hire local hunters to bring him a living specimen. The locals were hesitant, saying the Bei-Shung lived in very rugged terrain and were extremely difficult to find and kill, much less to try to capture one.
Father David took their reluctance as a ploy to extract more money from him, so he offered a handsome bonus. Chinese were loathe to give offense, so these agreed to take a crack at it.
In one of history’s great quirks of fate, after only twelve days the Chinese hunters returned with a living Bei-Shung they insisted had been secured only by astounding good fortune.
Their protestations seemed unlikely, but Father David didn’t care if they were embellishing the difficulty of their task. He knew their captive would make history worldwide, so he didn’t concern himself with the circumstances of its capture or the high price he had paid for it.
 |
For as great as Father David’s good fortune was with the capture, his luck soured in transporting his prize back to France. After a few days on a ship, the wild Bei-Shung became so agitated that it began ramming itself relentlessly against the cage bars, clearly intent on breaking free of captivity or dying in the effort to escape.
For humanitarian and moral reasons, Father David had no choice but to euthanize it. Having to kill his precious prize was a terrible blow to Father David, who found solace knowing he could still achieve his main goal of proving beyond doubt that the Bei-Shungs did exist.
If he could do that much by himself, he knew, well-equipped field teams would follow with sufficient methods of extracting the living Bei- Shungs they would no doubt easily capture.
He sent his “legend’s” preserved remains to a Paris Museum, creating a worldwide sensation.
Presaging what will probably happen after the first hominoid is officially presented in our own time, the world’s foremost scientific institutions in 1870 entered into a race to decide who would put the first living Bei-Shung on display.
They sent legions of topnotch experts — hunters, trappers, and scientists — swarming through the mountainous regions of Sichuan, a region the size of Arizona or Italy. All waited anxiously to see who would be the first to bring one in …
And they waited … and waited. By 1900 — 31 years since Father David’s find — all museums that financed expeditions had long since given up. By 1910, western media had renamed the Bei-Shungs Giant Pandas, and scientists had shoved them back into the mists of “legend.”
Despite Father David’s stuffed specimen being as real as it ever was, 41 years without so much as a follow-up sighting convinced most experts the panda was now extinct. Why? Because the men who had searched for it were all highly experienced woodsmen and skilled trappers.
 |
Cavorting around bamboo forests in Sichuan’s mountains looking for phantom pandas became a frontier jaunt for daring sportsmen with time and money to burn, like U.S. President Teddy Roosevelt’s two sons, Teddy, Jr., and Kermit.
In 1929, exactly 60 years after Father David’s misadventure, Teddy spied a panda in a tree and shot it, sending a bullet through that poor animal to puncture innumerable scientific egos, and giving it an iconic name — Teddy’s bear.
***
The rest of this story is fascinating, especially for women, but I’ll leave it for the eBook that deals only with hominoids. For now, let’s focus on what I said about the panda’s story being a perfect explanation for how hominoids manage to remain “undiscovered” in Western eyes.
Imagine 60 years of effort by top wilderness experts to secure one specimen of a numerous (at that time), distinctly marked, reasonably large, slow-moving, dimwitted animal existing in a rugged but confined area, living a daytime existence, and eating a highly restricted diet.
Consider how infinitely more difficult to locate and secure specimens of shrewdly intelligent, largely nocturnal, highly mobile omnivores that are absolute masters of enormous areas of some of the Earth’s most demanding environments.
Even utilizing the highest of today’s high-tech hunting and tracking equipment, it would be an exceptionally difficult task. But it can be, and someday it will be, done — on a regular basis. It will be the giant panda story all over again.
Intervention Theory suggests that Neanderthals are almas-kaptar type hominoids, the man-sized ones that dominate in eastern Europe. This does not mean they never migrated to other places in the world. Man-sized hominoids can be found living on every continent except Antarctica.
But, if the Neanderthals are almas that left no fossils accepted as such in the Americas, or in Africa or Australia, how can they be there now? It comes down to how death occurs in the wild.
Remember, the only known fossils of chimps are three teeth. Very few fossils of other higher primates exist, and all are questionable. This is because forests and jungles are the least likely environments to allow bones to fossilize.
Fossil bones are so rare because a very unique set of circumstances is needed to create them. First, a death must occur in such a way that the corpse — or some portion of it — will be covered over by sediments before scavengers scatter its bones, and bacteria and molds consume them.
Anaerobic (no oxygen) conditions are needed to neutralize all aerobic bacteria. These conditions are found in bogs, tarry pits, frozen tundra, etc.
Several death circumstances can create fossils:
- Burial in perpetually damp ground, where water can steadily leach minerals into the bone.
- Death in shallow water, with a body pressed into sediment by, say, a herd of hoofed animals at a kill site beside a muddy watering hole. The abdomen must be torn open so it doesn’t bloat.
- If the body floats, it snags on rocks or a limb so scavengers can’t reach it as it decays, and the body’s bones can drop into the sediment below.
- Death in a flash flood so the body is encased in enough damp soil to permit decaying of flesh and then leaching of minerals into the bones.
- Death in a cave, or outside it and the body is moved in. Numerous fossils are found in caves.
Once the death circumstances are ideal, during hundreds of years the minerals in sediment will replace the bones with precise stone replicas of themselves. Indeed, fossils are exact recastings of a corpse’s bones into mineralized stone.
As fossilization occurs, more sediment piles on top of the fossil while the land it has become a part of shifts, twists, buckles, or folds as plate tectonics constantly reshapes the planet’s crust.
Those movements — along with weathering or mining or other excavations — occasionally will expose fossils to view, after several thousand years or several millions. Thus, the creation of, and discovery of, any fossil is the end result of improbabilities stacked upon improbabilities.
For obvious reasons, fossilization occurs most often among aquatic (water-dwelling) animals. The rate drops significantly among terrestrial (land-dwelling) animals, but a fair number are still found because they tend to visit water holes daily, and water holes are where a great deal of hunting is done by Earth’s natural predators.
Among arboreal (forest dwelling) animals, the fossilization rate is so very low because they normally obtain their liquids from fruits and such within the habitat, relieving them of the need to routinely risk their lives at water holes.
This explains why all pre-human fossils are so rare. They all live in forested areas, and when they die — in whatever circumstances — their bones dissolve to nothing in only a few weeks.
With that said, why are Neanderthals far and away the most commonly found pre-human fossils? Because many places in Europe have caves, which are ideal homes for hominoids when Ice Ages make living outside difficult.
This is not to suggest Neanderthals weren’t fully hair-covered and able to withstand sub- zero temperatures. However, they weren’t stupid, either, so in Ice Age Europe if they found a cave to live in, why wouldn’t they?
The bottom line is this: If it can be shown that Neanderthals were hominoids, that means they were — and remain — descendants of creatures that lived and have endured since the Miocene.
Hominoids are one of two “fantasy” creatures that can dismantle Evolutionary Theory in one fell swoop (off-world aliens are the other).
The day a hominoid is presented to the world for analysis — dead or alive — is the day science can no longer prop up its evolutionary dogma.
When macroevolution clatters into history’s dustbin, Divine Creation and Intelligent Design will follow right behind it. Intervention Theory will become the most plausible explanation for how humans and all life came to be on Earth.
In the end, Intragalactic Terraformers — which now might seem absurd to many people — will become the choice everyone settles on because anything else will seem laughably implausible.
In the interim, until undeniable proof is in hand, our only course of action is to focus on the best evidence for hominoid reality; and that, in my opinion, is most strongly seen in Neanderthals.
For 100 years mainstream scientists insisted Neanderthals led to Cro-Magnons in a direct line, even though not nearly enough time was available to account for the numerous physical differences by means of Darwin’s gradualism.
That much-too-narrow time gap between them became home to the “missing link,” a mythical fossil pre-human that, when found, would show the “transition” from one species to the other.
Since the late 1990s, when it became clear that a Neanderthal-human link was tenuous at best, the mainstream grudgingly shifted their focus to Erectus/Ergaster to establish a new “transition” species from which humans must have evolved.
Now mainstreamers consider Neanderthals a “dead end” on an “evolutionary path” that ends 28,000 years ago. They refuse to consider any evidence that contradicts the idea of evolution being life’s bottom line. Their dogma prevails.
Despite their resistance, they cannot make that evidence disappear, so let’s consider some of it.
 |
 |
 |
In 1978, near a place called Laetoli in northern Tanzania, East Africa, a long trail of fossilized hominid footprints were discovered. They dated to 3.7 mya, which made them contemporaneous with early the Australopithecines, like Lucy.
Those tracks were laid down on a plain near a volcano spewing layers of muddy ashfall. After one eruption deposited a pristine layer, at least two early pre-humans — one smaller than the other — strolled side-by-side across the ashfall.
Those tracks dried to form solid impressions of the feet that made them, then more ash covered them, perfectly preserving them until one was accidentally discovered and the others revealed.
Their foot sizes were 7.3" and 8.5" respectively, making them about 4 to 5 feet tall. The smaller one left several excellent prints, which leave no doubt that both were bipedal in gait and stride.
 |
 |
 |
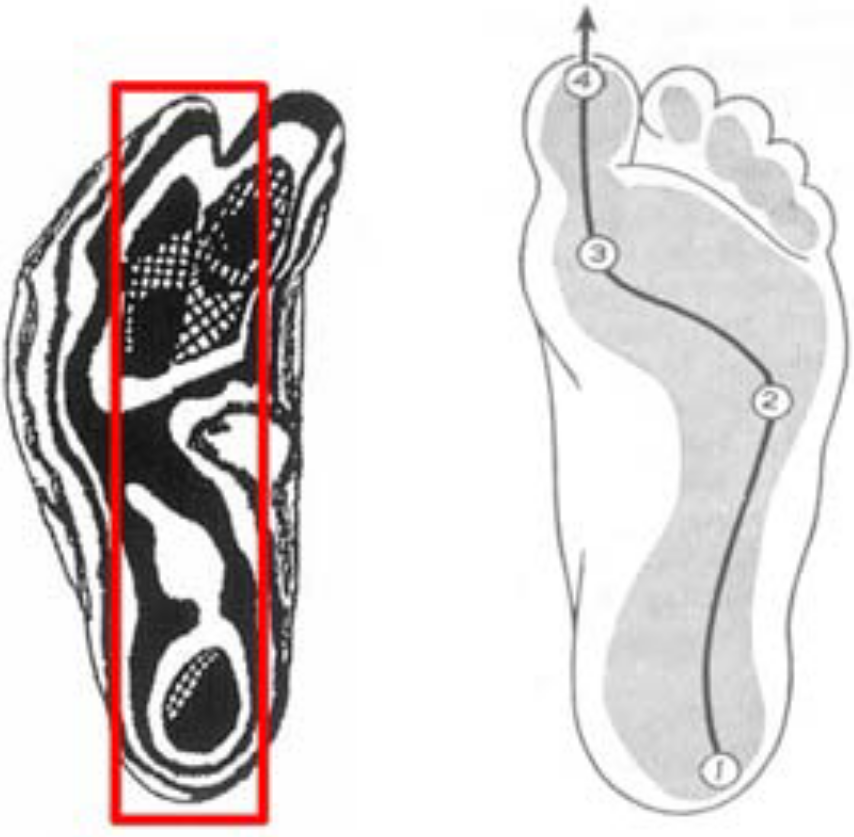 |
 |
Despite undoubted bipedality, their feet are not like human feet. They have a shallow, barely discernable arch, but a significantly different length-to-width ratio. Their small toes have different shapes, alignments, and functions.
Even greater difference is seen in their big toes, which have an inward, pigeon-toed slant. But, the most significant differences are in how they operate biomechanically. Their stride is unique.
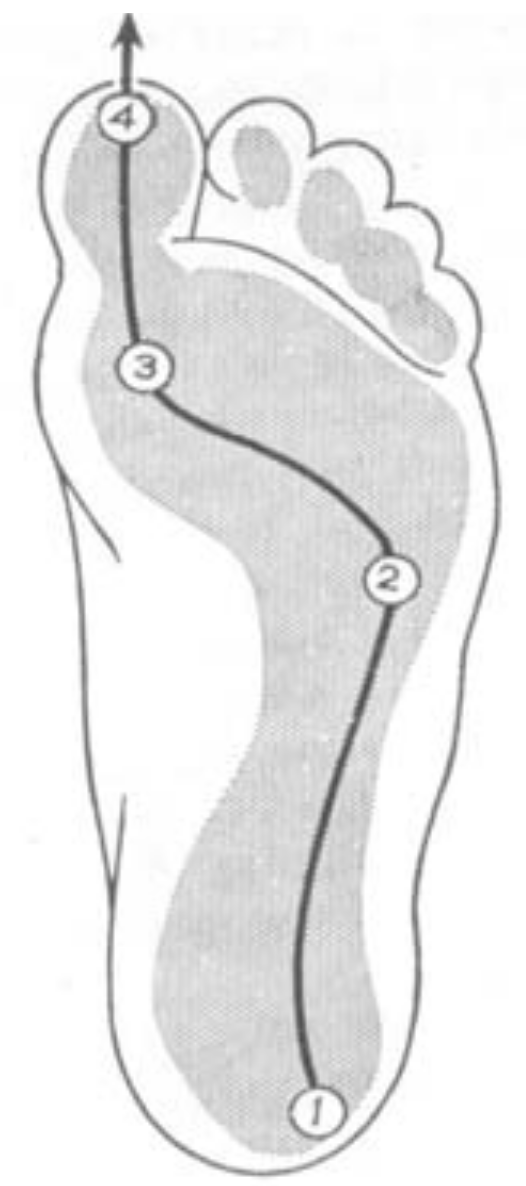 |
The top right photo is of a human foot toeing off for a stride. As a step proceeds, our momentum first strikes the heel (1), which “plants” us and pushes our weight forward, transferring it along the outer edge of our foot sole, skirting the arch area until our weight is directly over the ankle.
Once the midpoint of any stride is reached and our weight passes beyond the arch area, it does a sharp crossover (2) into the “ball” of the foot.
The ball (3) transfers our weight forward and out along the big toe (4), and to a much lesser extent to the small toes, which act as balancers for the step thrust generated by the big toe.
2nd from top, is a pressure-sensitive analysis of the Laetoli foot. Notice how much it differs from the human foot. Notice especially how much the ankle positions differ (see below).
When walking, humans generate a heavy heel strike followed by a hard push-off through the ball of the foot and toes, especially the big toe.
In stark contrast, the Laetoli footfall is well balanced from rear to front, with a light heel strike and the whole forefoot used to stride off, with all five toes providing propulsive force.
What this means is that the Australopithecines of 4 mya to 2 mya walked far more efficiently than we humans walk. Slow-motion analysis of our walk shows how discombobulated we are.
Each step begins with a jarring heel strike, our momentum swings around our arch and into the ball, then thrusts out through the big toe. In the middle of that, we lock our knee and put torque on our hip joint as it carries our trunk forward.
It is no wonder our leg joints decay as we age. Laetoli feet work with a more stable plane than humans, impacting the heel, flattened arch, and forefeet about equally, giving them a smoother, better balanced weight distribution than us.
The entire surface of their feet glides down to a landing, then their weight shifts forward along the foot’s midline, meaning no locked knees and a smoother “carry” through each step.
The weight shift is a gentle “S” along the foot’s midline, eliminating our jerky motion. Then the push-off comes from just behind their toes — the big toe and the small ones working in concert — rather than a human’s push-off with the big toe.
Such a smooth, gliding walk could only come from a biped keeping its knees semi-bent with each step, never fully locking up at any point.
It’s also using its thighs — bones and muscles — as shock absorbers and weight carriers rather than — as with humans — letting the knee joints and hip joints absorb the stresses of each step.
So, this bent-kneed, even-keeled, well-balanced gait tells us the two Australopithecines walking across that stretch of volcanic ash 3.7 mya were utilizing bipedality in its most efficient form. But, as always, the mainstream begs to differ.
They try … ohhhhh, how they try. Remember, the mainstream’s purpose is to present facts and figures and images that, on cursory inspection by the uninformed, will seem logical and true.
This is how their brainwashing programs work, so I will pick apart this one example because it is so important to Intervention Theory, and it is so egregious in its misuse of their “authority.”
What we will discuss is a peer-reviewed (don’t get me started on the massive corruption of that absurd process) paper about the Laetoli tracks that can be looked up by anyone so inclined.
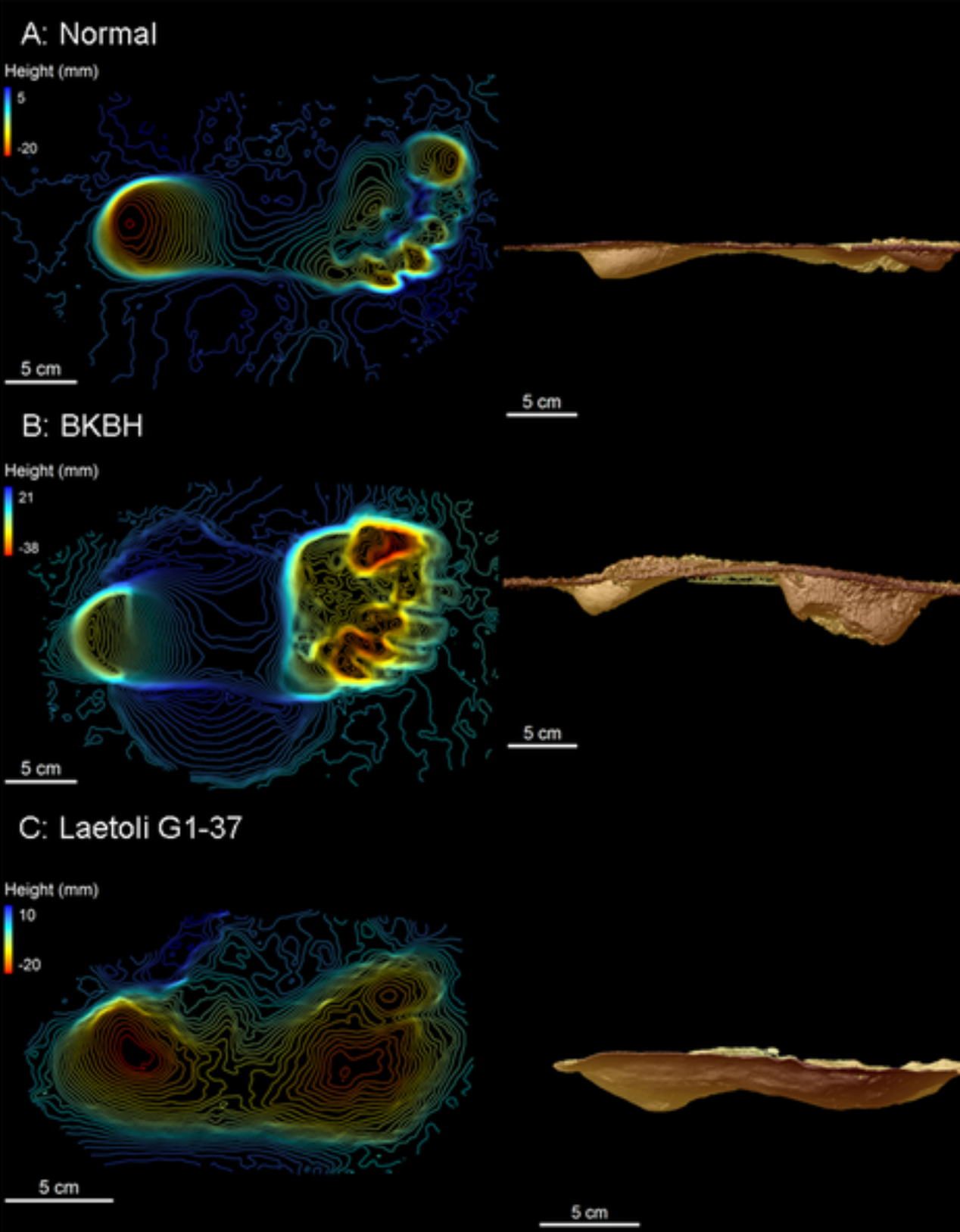 |
The paper analyzes the results of an experiment comparing the Laetoli tracks with two sets of human tracks in sand — one walking normally, and one with what the researchers refer to as a “bent-knee, bent-hip” (BKBH) technique.
To the right, pressure points are digitally expressed as areas of “light.” On the right are sidelong views of each step’s depth and shape.
At the top, a human walking normally shows much more light in the heel and toe areas than the Laetoli track at bottom, which has a very “even” dispersal of “light” in every respect.
The increased depth of the Laetoli track is explained by the inability of the researchers to match the damp sand they used to the damp ash that made the Laetoli prints. We might wonder why sand was used instead of ash, but we have to assume the researchers had a reason for it.
According to the experiment results, the human track and the Laetoli track produced about the same heel depth and forefoot depth. Yet their pressure images are clear that the human heel strikes with more force, and the forefoot pushes off with more force than did the Laetoli foot.
I have already discussed the smooth, uniform prints from Laetoli, and the pressure image in this experiment clearly bears that out. It shows remarkably even pressure all across the print, with virtually no “light” anywhere on the track.
Despite the human print being so obviously different from the Laetoli print, the researchers assure us they are such a close match that they “prove” the Laetoli track makers walked very much like humans walk today. Do you buy it?
If this were a legitimate comparison, there is no way the Laetoli track could produce the same depth of imprint at the heel and forefoot as the human did. This is a completely bogus finding.
So, right out of the box the experimenters are “fudging” their results. And why? Because it is important for Laetoli to be pre-human in order to prop up the “evolution-at-all-costs” scenario.
***
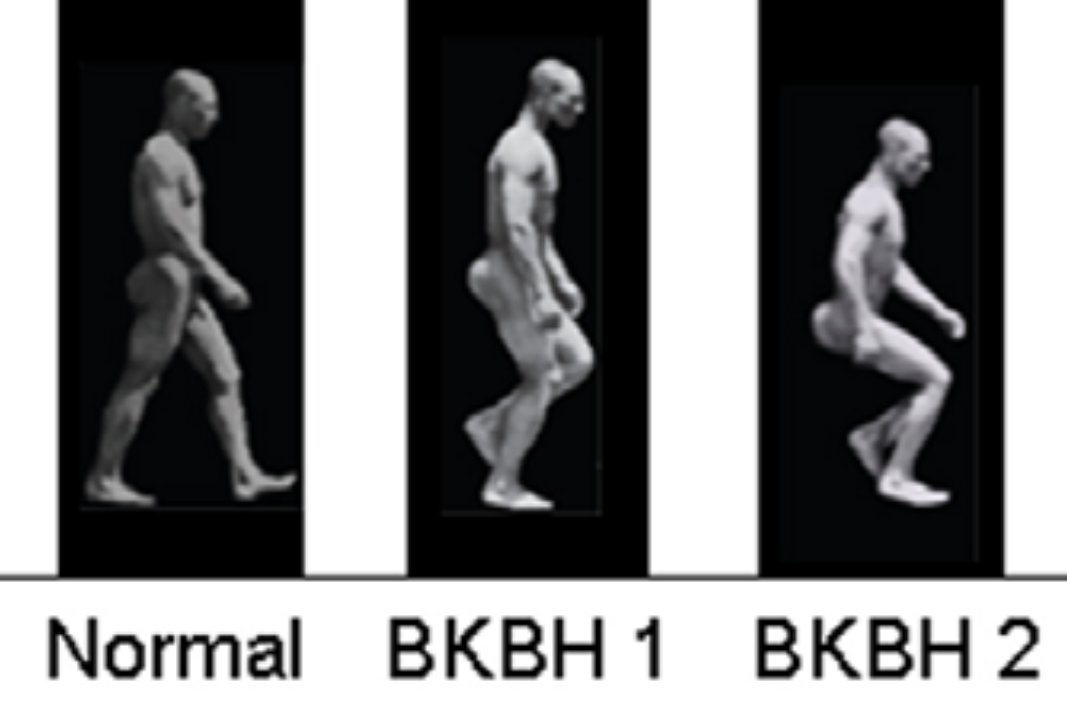 |
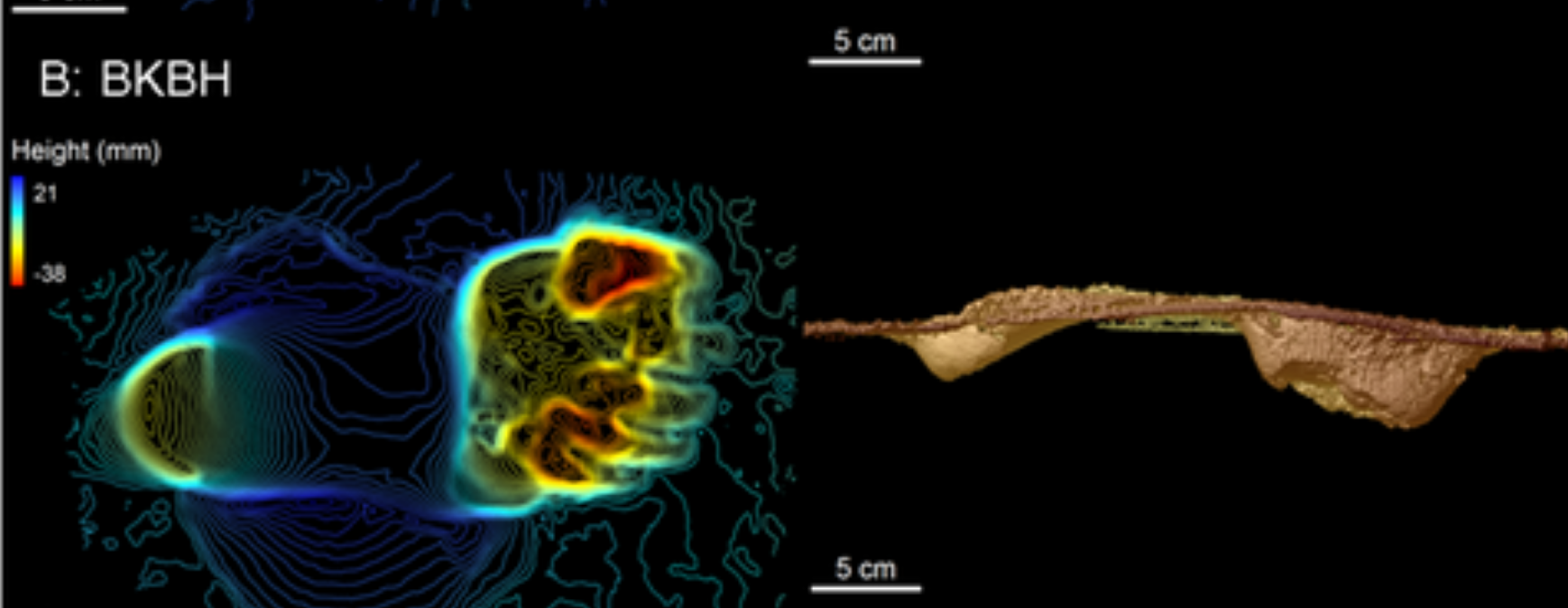 |
Another egregious problem with the experiment is the middle illustration (right) created when subjects walked with the BKBH technique of primates.
In BKBH1, the subjects walked with slightly bent knees in a “Groucho Marx” style, as I’m certain all pre-humans/hominoids walked.
In BKBH2, the subjects bent well over, with their knees bent rather drastically, imitating the “knuckle-dragging” primate walk rather than the smooth striding “Groucho Marx” walk.
In the graphic with the BKBH comparison in its middle, they don’t say which one we’re shown, but we can confidently suggest it was BKBH2, because that shifts a body’s center of gravity so far forward that it falls onto the ball of the foot.
This is most definitely not the track of someone walking with the “gliding” stride of hominoids.
Anyone living near a sandy beach or strand can check this. Walk barefoot on damp sand. Walk normally, then try to “glide” like a hominoid, then drop your hands down like a “knuckle dragger.” The differences will be obvious.
Your “gliding” track will resemble the Laetoli track much more than your “normal” track or your knuckle-dragging track. And when you do that, you will see how this report is designed to mislead people into believing the makers of the Laetoli tracks were, at 3.7 bya, nearly human.
One last criticism of the experiment is that humans and the Australopithecines that made the Laetoli trackway are physiologically like cats and dogs. Their knees don’t extend straight the way ours do, and our knees and feet can’t begin to function in the ways that theirs do.
The bottom line is that this experiment was never a good one, and was never intended to be a good one. It was meant to serve the purpose of bolstering the mainstream dogma that all of the early bipeds had to be incipient humans.
Nothing could be further from the truth.
 |
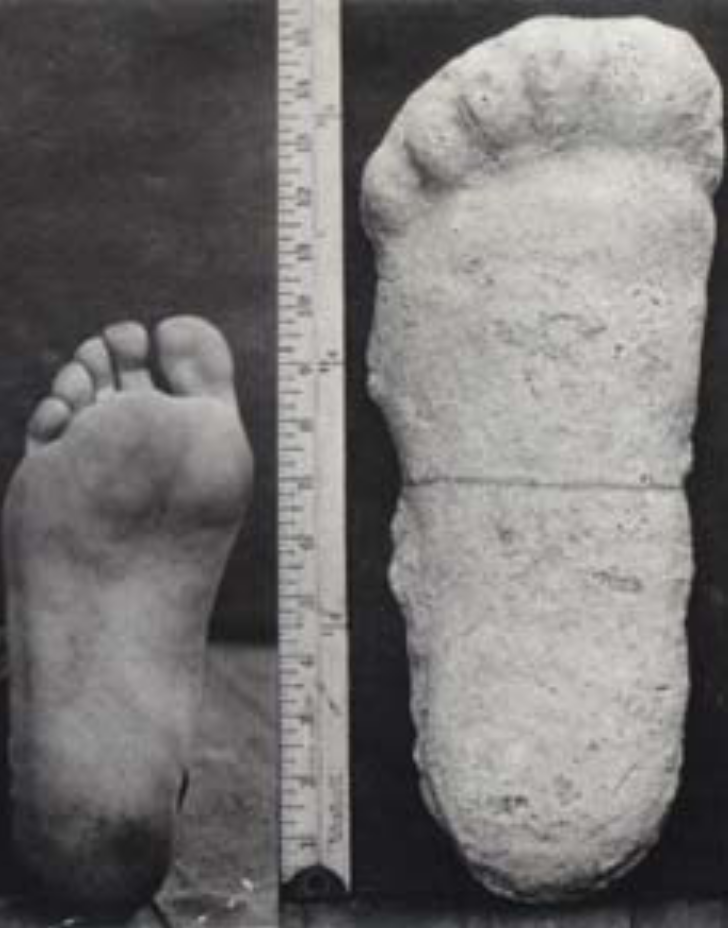 |
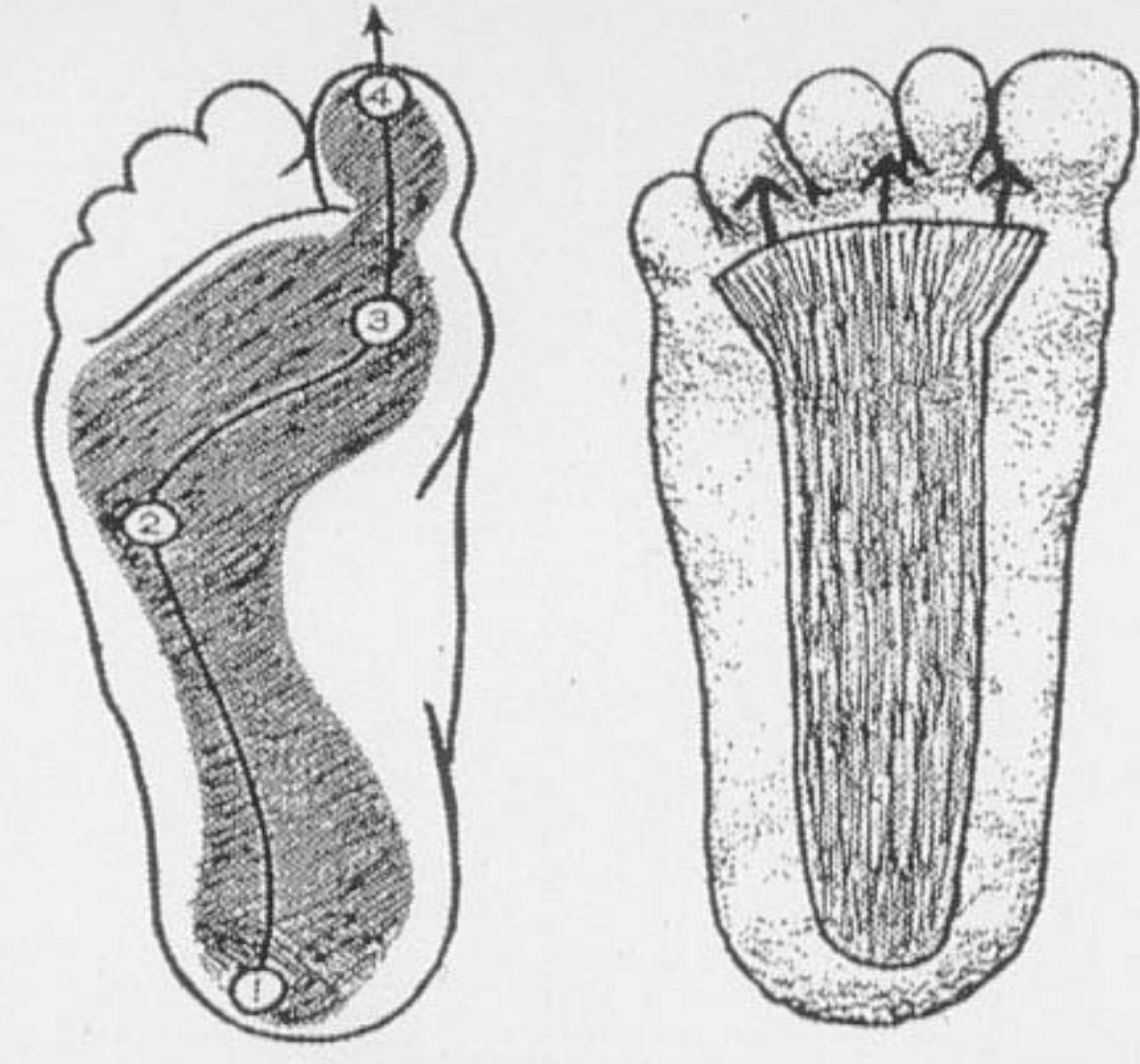 |
That’s right, Patty in the Patterson-Gimlin film. She walked exactly like the Laetoli tracks say those creatures walked. And Patty’s feet look and worked the same when she left her tracks.
That is Rene Dahinden’s foot at left (top right), which is three inches longer than the largest of the Laetoli track makers, beside one of Patty’s tracks on the right. The lines of motion as both feet walk are seen in the diagram (below right).
The amazing truth that mainstream scientists never want to admit or to advertise in any way, is that those Australopithecine track makers at Laetoli, though nothing like humans other than sharing a mode of locomotion, at 3.7 mya had already perfected upright walking to a degree humans haven’t yet mastered — and never will.
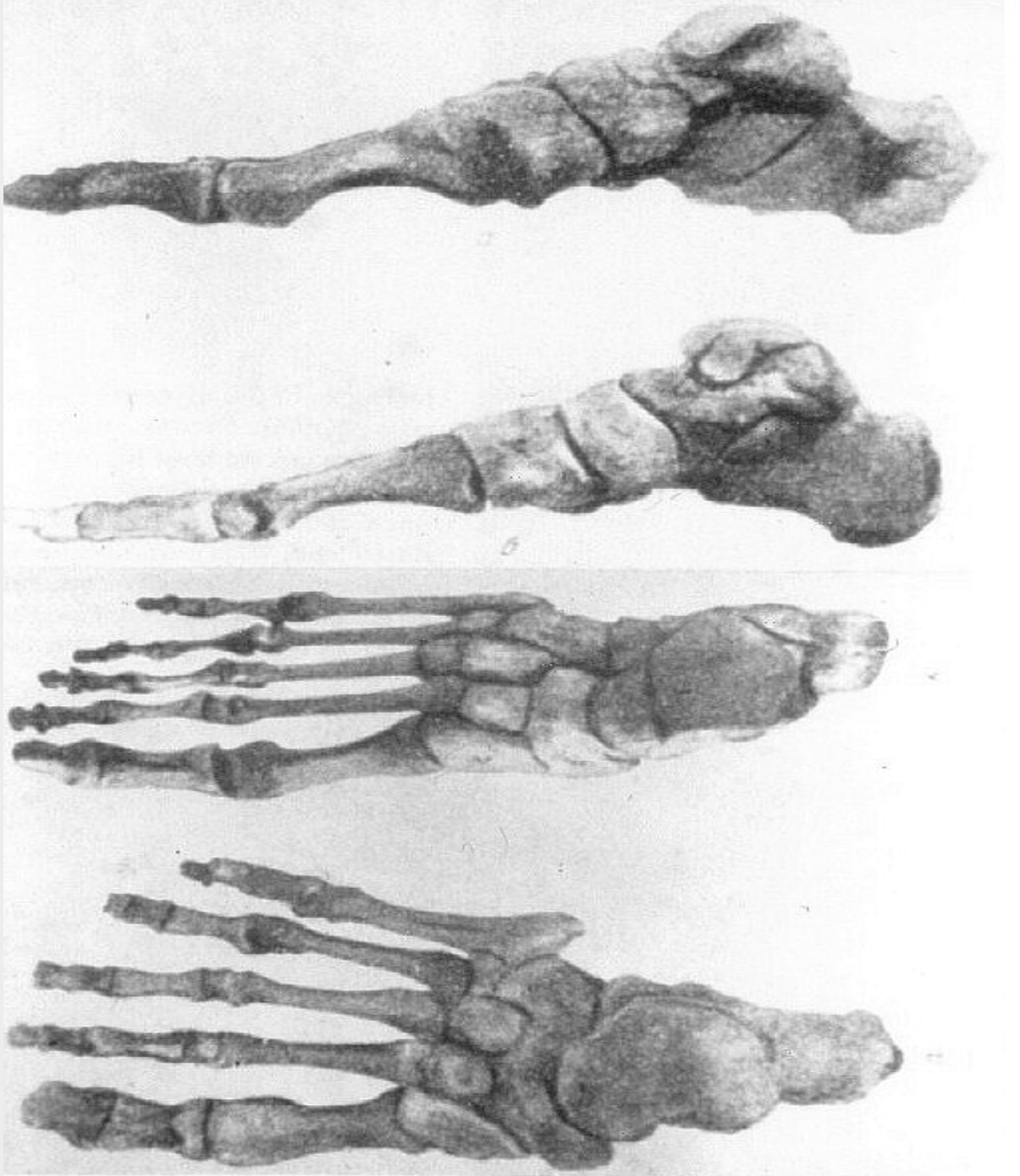 |
When it comes to tracks and making them, Neanderthals are in a class by themselves. While we do not have film of them walking (Patty was a bigfoot/sasquatch type), we do have the next best thing — their actual feet!
Right, Neanderthal feet bracket human feet at top and bottom, the human pair in the middle.
It takes only one glance to see the huge range of differences between them. The bulk of the bone is obvious in the Neanderthals, as is the extra length of the heel. Notice how much larger the ankle bone is, and how much farther forward.
Notice at bottom the squat splay of the mid-foot bones, the relative sameness of size in the toes. See the human’s zig-zag from heel to midfoot, due to the momentum swinging through them.
The Neanderthal foot is wide, broad, flat, and rock solid, exactly like the Australopithecine foot 3 million years earlier. The only change in that whole time was growing 3" to 4" longer.
This is not a bigfoot or an almas foot. But when one of them is produced — and eventually it will happen — it will be a replica of the foot above.
Why? Because all hominoids are basically the same kind of creatures, and those creatures are the several bipedal apes that walked out of the Miocene epoch starting 20+ million years ago.
***
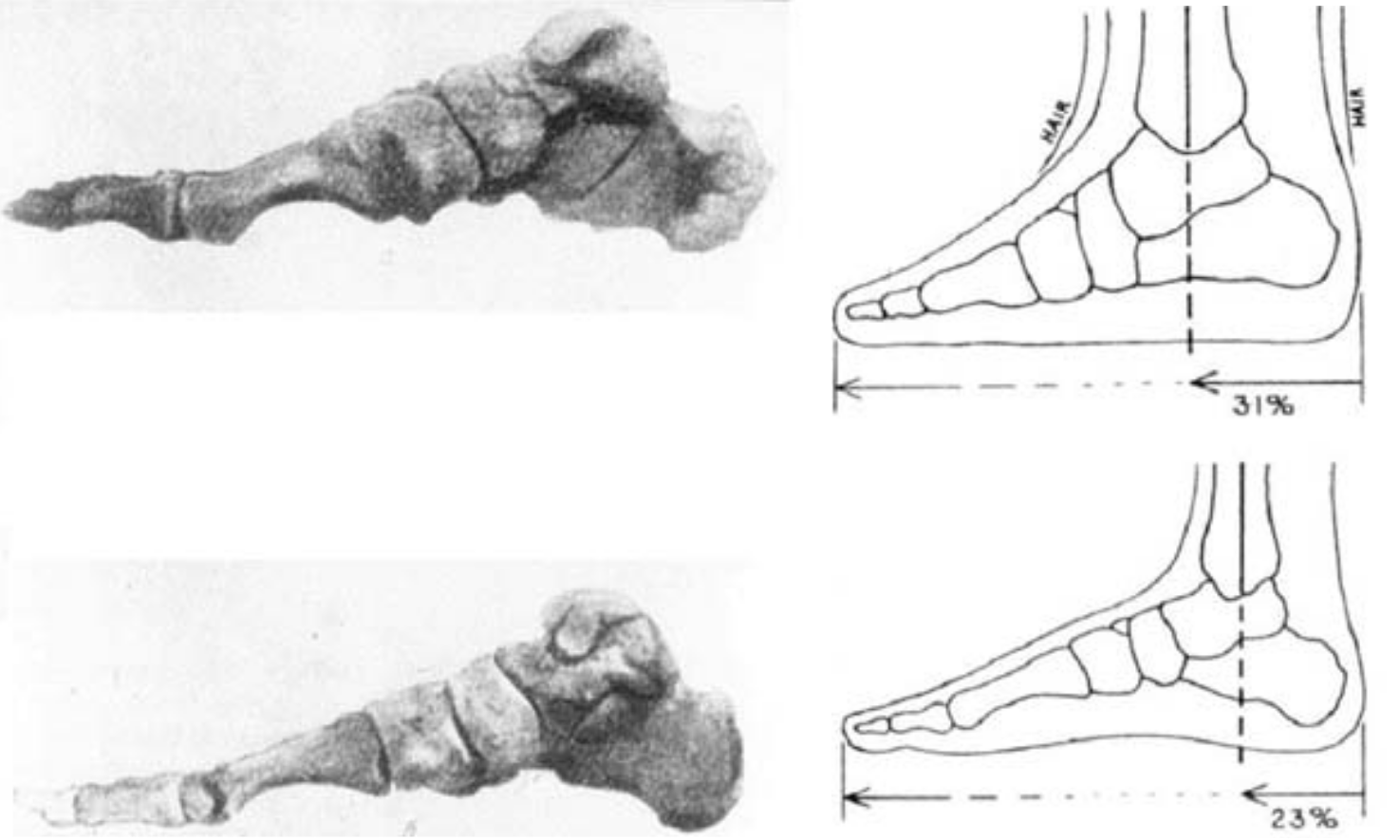 |
Even though we don’t have a bigfoot foot, we do have Dr. Grover Krantz’s drawing of Patty’s foot anatomy based on careful observation and measurements of her feet and ankles in the film.
Once again, we have a striking resemblance between Neanderthal and bigfoot, but striking dissimilarity between them and a human.
How many of these comparisons do you think mainstream science would have to see to take a step back and say, “Welllll, maybe there’s a bit more to this cockamamie Intervention Theory than appears at first glance. Maybe we need to rethink our position on it. Maybe we’re wrong.”
***
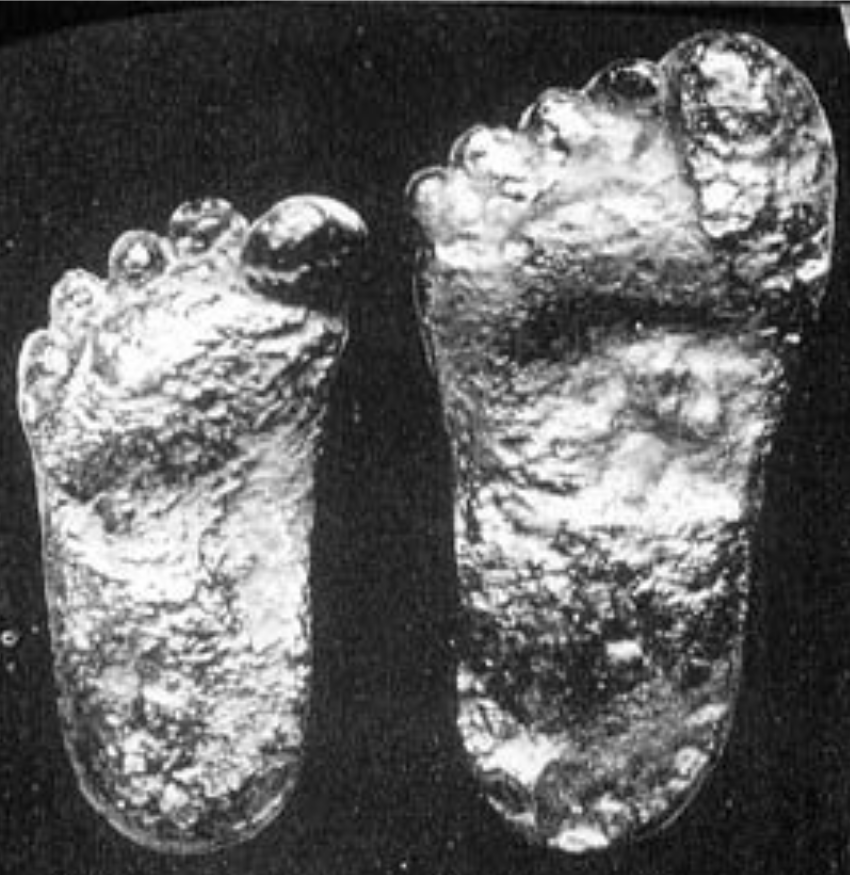 |
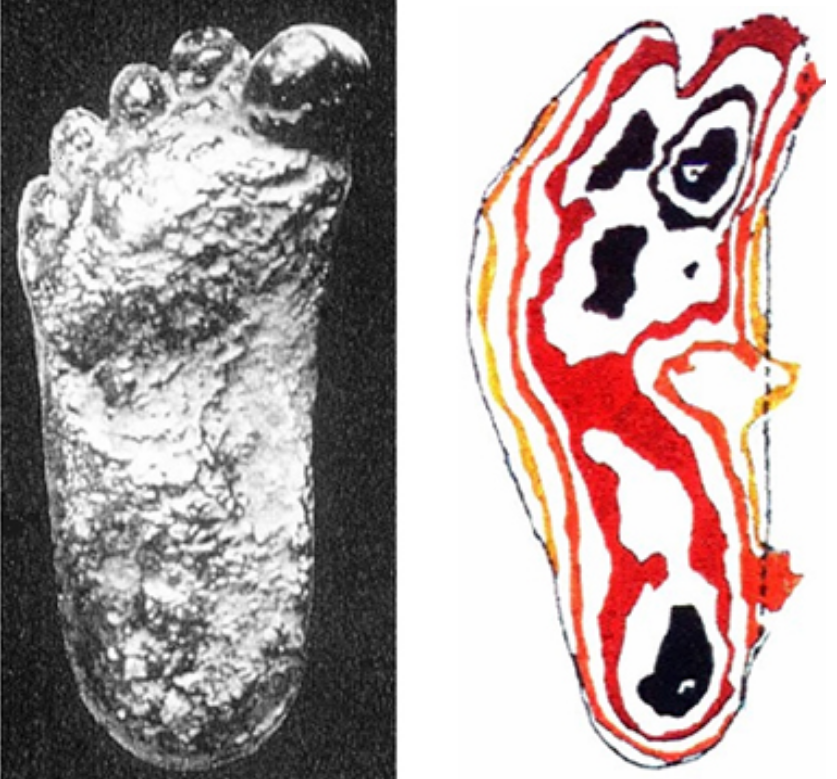 |
 |
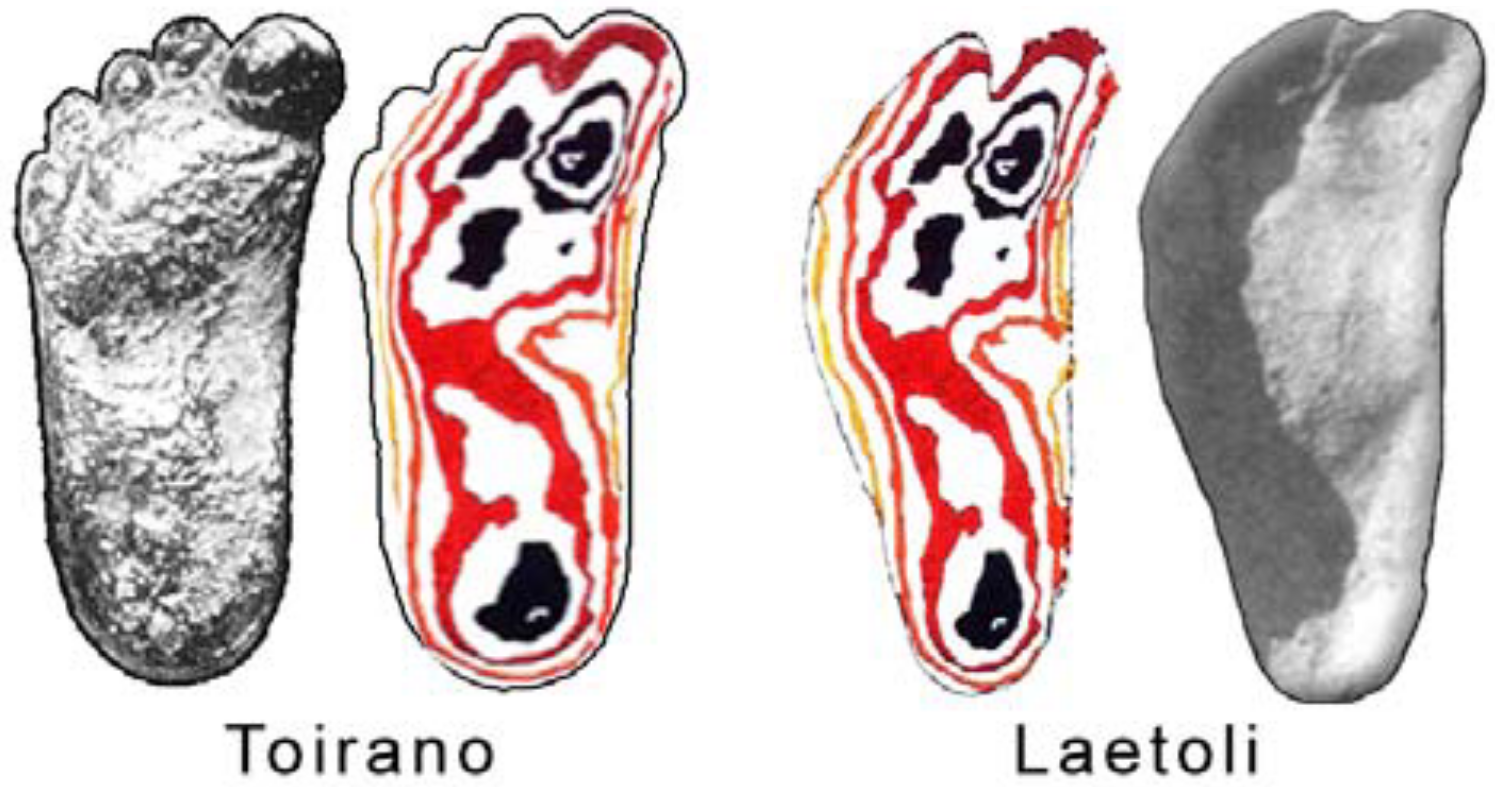 |
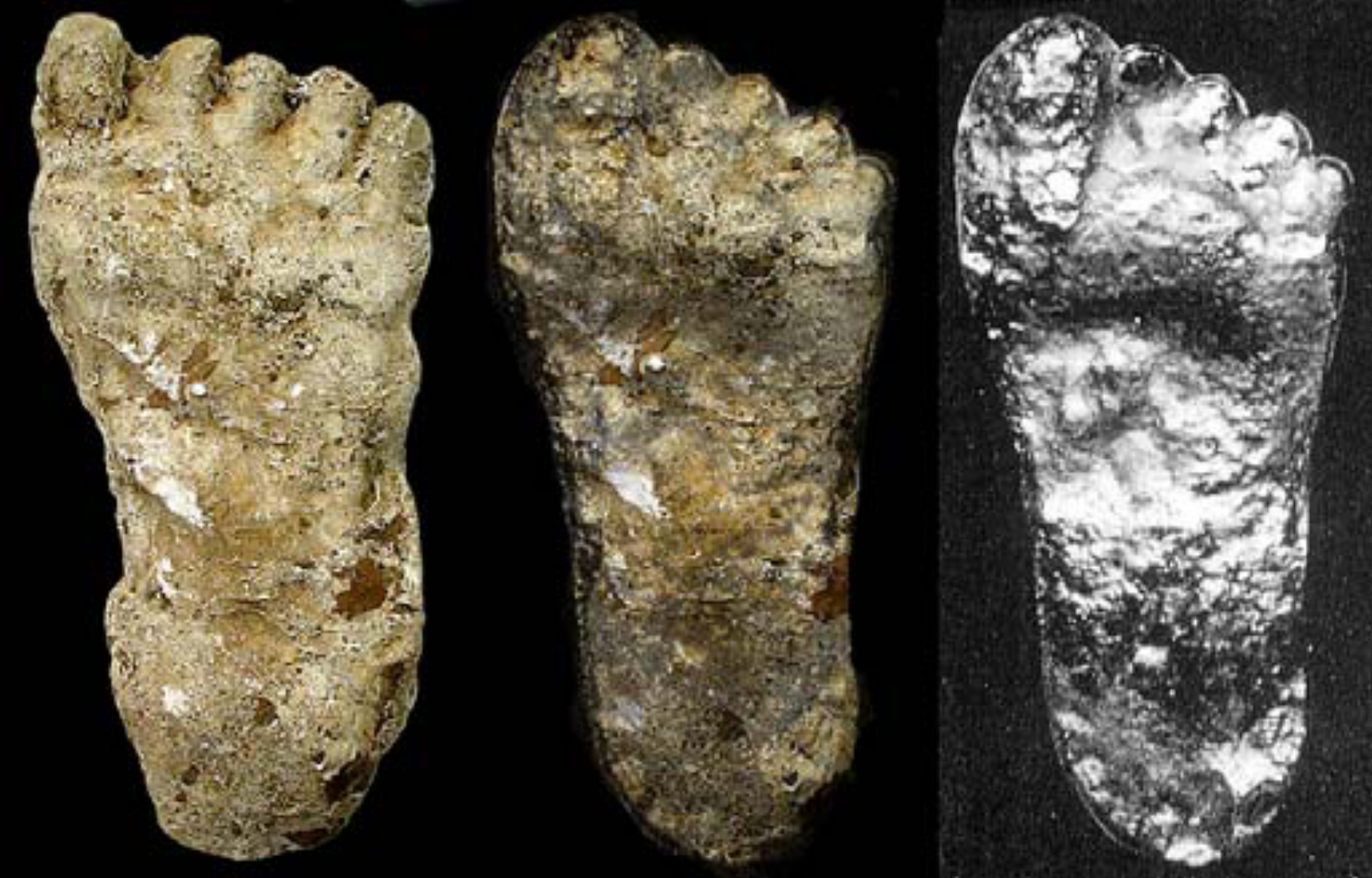 |
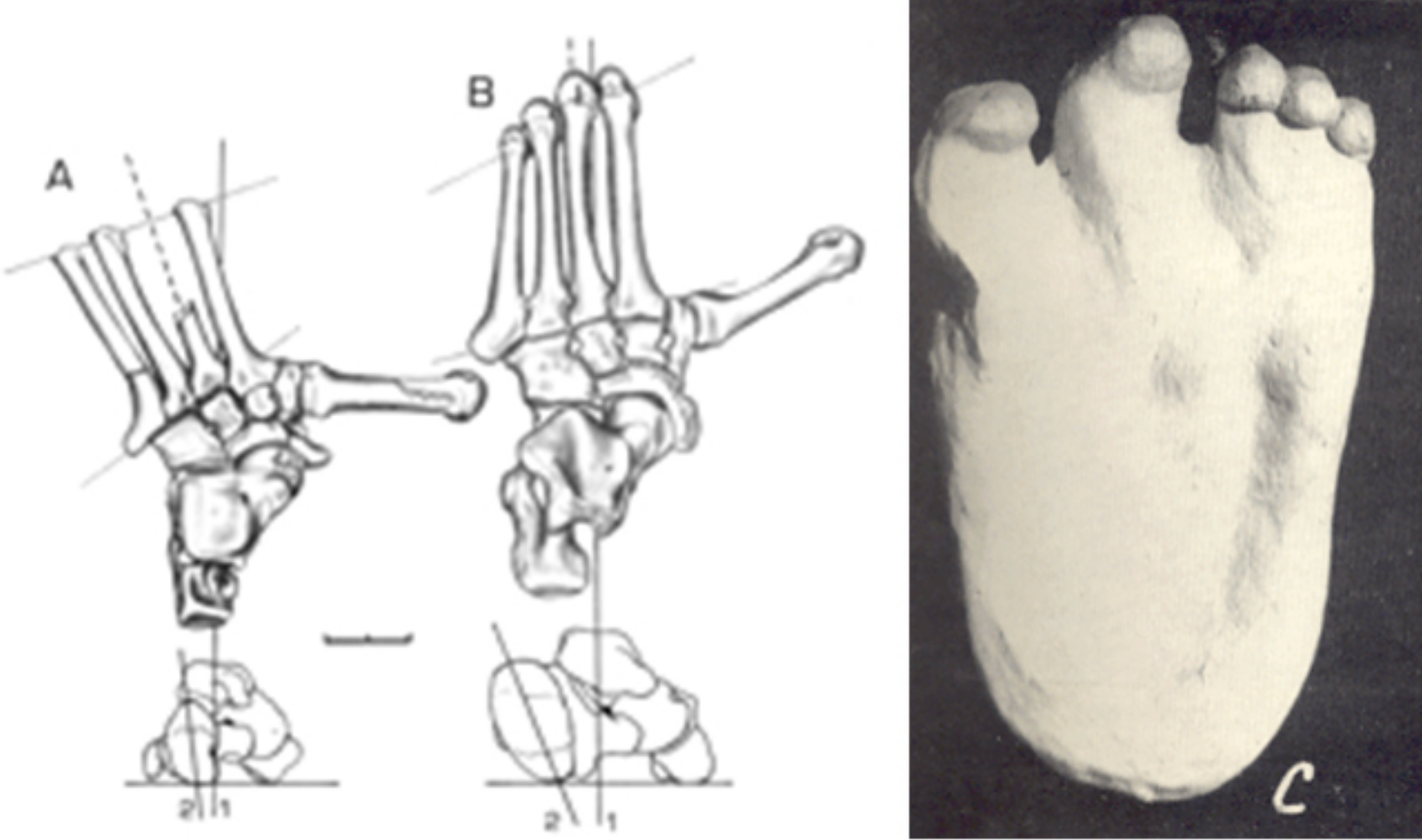 |
Top right is another set of Australopithecine tracks, not as famous as the Laetoli tracks but, as can easily be seen, these share a few similarities. The major difference is that Laetoli is a track and these are the mirror-image casts of tracks.
The cast at the left is considered a female and the one at right, a male. This is the same basic configuration of the tracks at Laetoli 3.7 mya.
Both are assumed to be made by females. The toes have the same degree of slant, the “push off” area is clearly the entire forefoot, the ankle is in the same place, but the heel is wider.
This can be due to variations in print mediums, because one of the other Laetoli prints shows a heel that the cast fits perfectly (3rd from top).
Now that we have that similarity established, it’s time to confess that I have inveigled you.
Inveigling is “to lead astray artfully; to deceive with style or class.” I prefer that to lying. But, in fact, the “new” Australo is a Neanderthal!
Those new tracks (3rd from top) came from the clay floor of a cave in Toirano, Italy, where many others like it were found. They were created in softened clay, which does not give up dates well, so the time they were laid down can only be approximated.
Bear bones found in the cave have been dated to 25,000 years ago. We can assume the bears and Neanderthals did not share the cave, so we further assume above or below 25,000. Since no Neanderthal bones are known to have been found after 25,000 years, the best guess for the tracks would be prior to 25,000 years ago.
4th from top are both tracks, with the Toirano pressure imprint extrapolated from the Laetoli imprint. Recall that Toirano is a cast from a solidified track, while Laetoli is a solidified track.
Both Toirano, at maybe 30,000 years ago, and Laetoli, definitely from 3.7 million years ago, mean that the feet propelling Australopithecines like Lucy microevolved relatively little in close to 4 million years to become Neanderthal feet!
It also means (to anyone with an open mind, of course, which doesn’t include mainstreamers), that every one of the so-called pre-humans that followed the Australopithecines, would have walked on a foot with the same biomechanics.
5th from top is one of Patty’s tracks, and in the middle is the male foot at Toirano adjusted to its size and coloring. Amazingly similar, eh?
(bottom right) Remember, at 9 mya Oreopithecus walked on a highly unusual foot (left), and the yeti/ abominable snowman type hominoid (right) also walks on a highly unusual foot. However, we can say with confidence that no hominoid walks on a foot closely similar to humans.
The first truly human foot does not have a clear arrival point until Cro-Magnons around 60,000 years ago. The foot bones of every pre-human before them is in one way or another debatable.
[Among scavengers, the relatively hairless feet and hands of bipedal prey seem to have been four cherries on top of those skin bags of meat, so they were gobbled first and thus have always been difficult for archeologists to recover.]
***
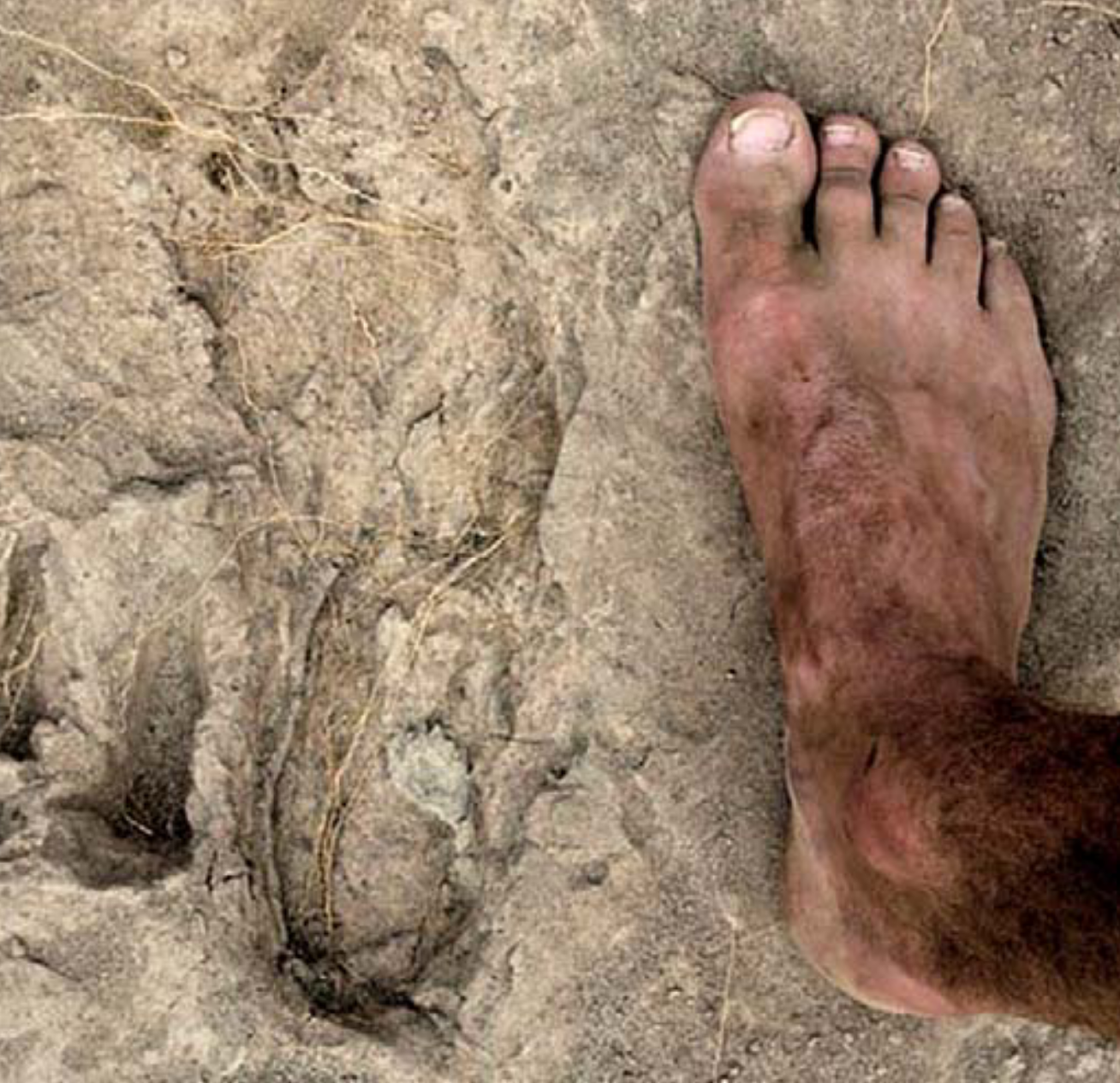 |
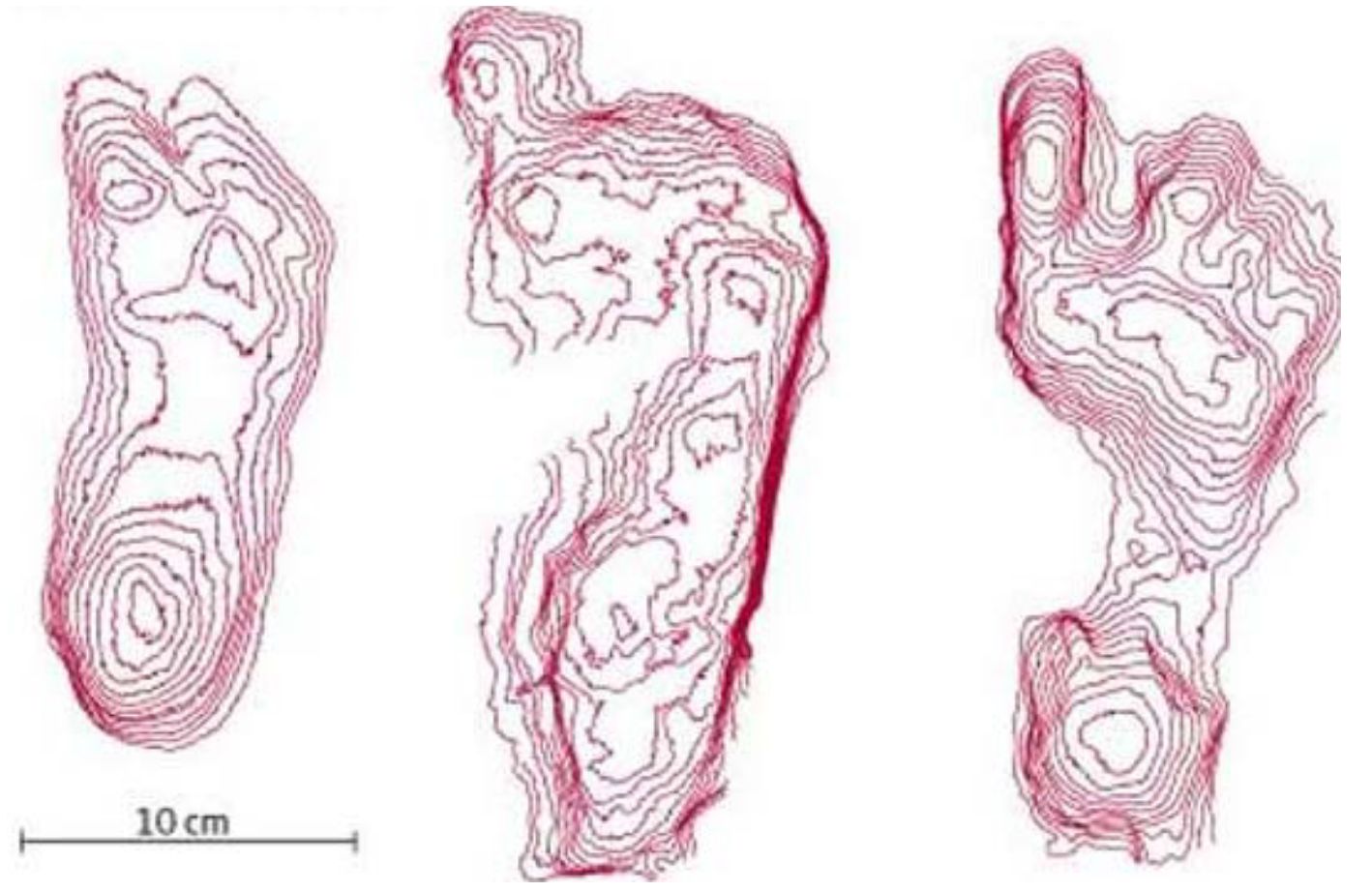 |
While discussing feet, we must mention two lines of tracks laid down near Ileret, Kenya, by Homo Erectus, 1.5 mya. These are the second- oldest, after Laetoli. (top right} a print is compared to a human foot that wears a size 9 (US) shoe.
Mainstream science considers this track and the others with it to be all but human, because that is what they need it to be to prop up evolution.
In another pressure-sensitive reconstruction (bottom right), we see the Ileret track (at center) shapes up as more like the Laetoli track (left) than a human (right).
No matter how the mainstream tries to bluff and bluster about the feet issues, inevitably the truth will come out. We don’t find human-like feet until real humans appear with Cro-Magnons.
Everything prior — including Neanderthals — is something else, which is bipedal Miocene apes, and several of those exist today as hominoids.
 |
 |
 |
 |
 |
Neanderthal hands were also distinctly different from humans. Compared top right are the tips of forefingers from a Neanderthal and a human.
That clearly shows how brutishly strong they were compared to us. Now take a look at a comparison of their thigh bones and ours.
Bone thickness directly correlates with physical strength. A bone’s thickness is determined by the torque that can be applied to it by muscles that attach to it. With bones like Neanderthals, their strength and power would be difficult for us to imagine, and impossible to duplicate.
Now, what about hands? 3rd from top is one from a Cro-Magnon at left, and Neanderthal at right.
Once again, the thickness between them is stark enough to be convincing that no test of strength would end in favor of a Cro-Magnon (human).
4th from top is the hand of an Australopithecus Sediba, the recently discovered pre-humans dating to 1.9 mya. Notice how it compares to a Neanderthal hand of around 30,000 years ago.
As with the Laetoli tracks 3.7 mya, Sediba’s hand is more human-like than ape-like, yet it has clear ape-like parts, including long fingers and a thumb curving out from the hand rather than the human thumb’s straight-line angle.
Does that unusually curved thumb strike any memory chords with you? Think back …
That’s the Minnesota Iceman sporting the same kind of hand and thumb alignment found in the hands of Neanderthals. Sooooo … could it be possible that Neanderthals were merely one of the almas/kaptar type of hominoids alive today? Naaaah! No way! Gotta be just a coincidence!
Also, it just so happens that a thumb like the one carried by the Iceman and Neanderthals and Sediba would be useful for very precise degrees of manipulation with the forefinger, such as picking berries or plucking body lice.
However, unless their thumb pads (the lump of muscle at the base of human thumbs) were far larger than the Iceman’s, their thumbs would not be useful for the “power” grip required to grasp and effectively wield tools or weapons.
Thus, using an axe or throwing a spear would probably be hard, if not impossible, for them. However, using hand tools such as the flaked stone tools in use for millions of years, would be perfect for them. Yet another coincidence!
***
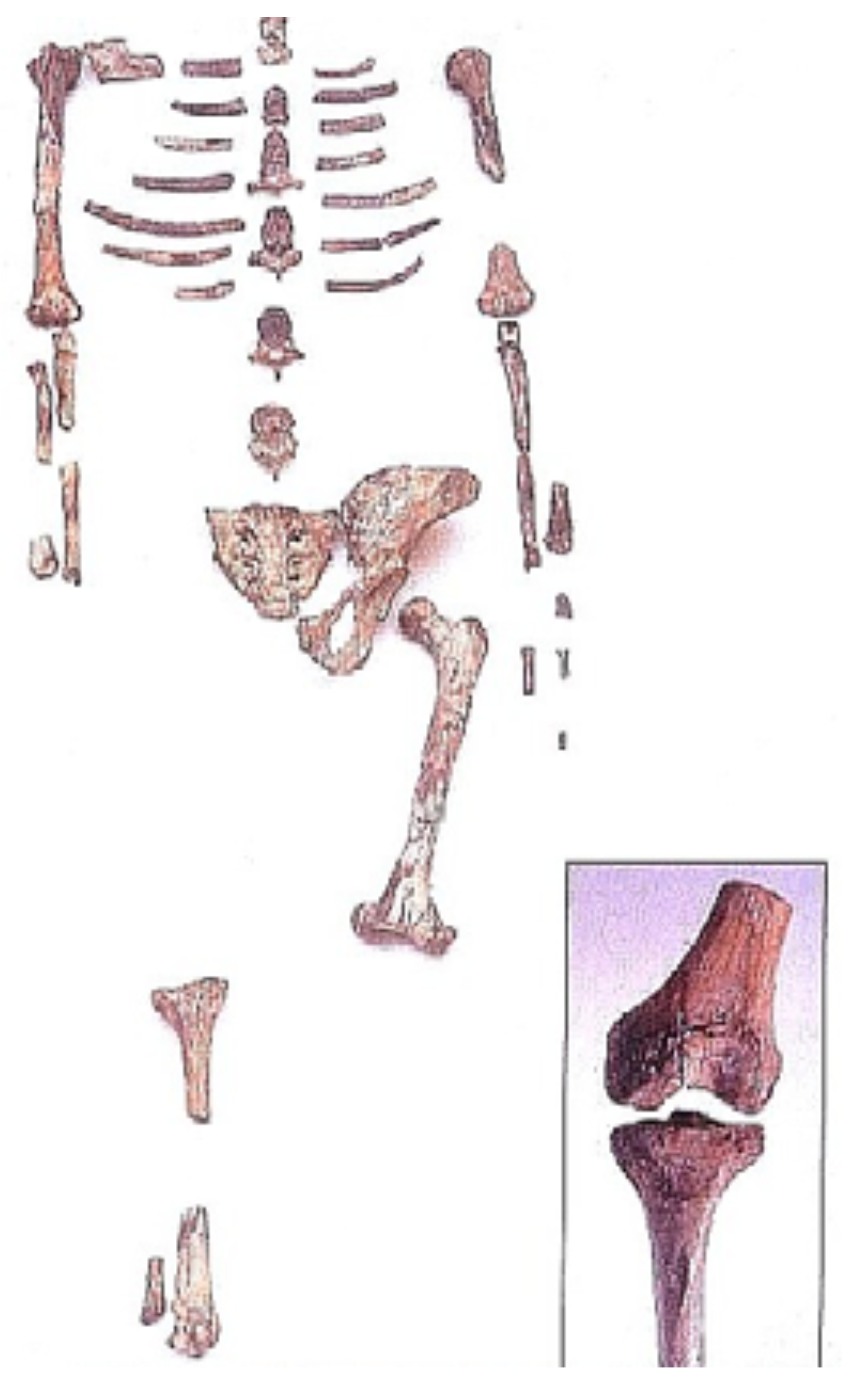 |
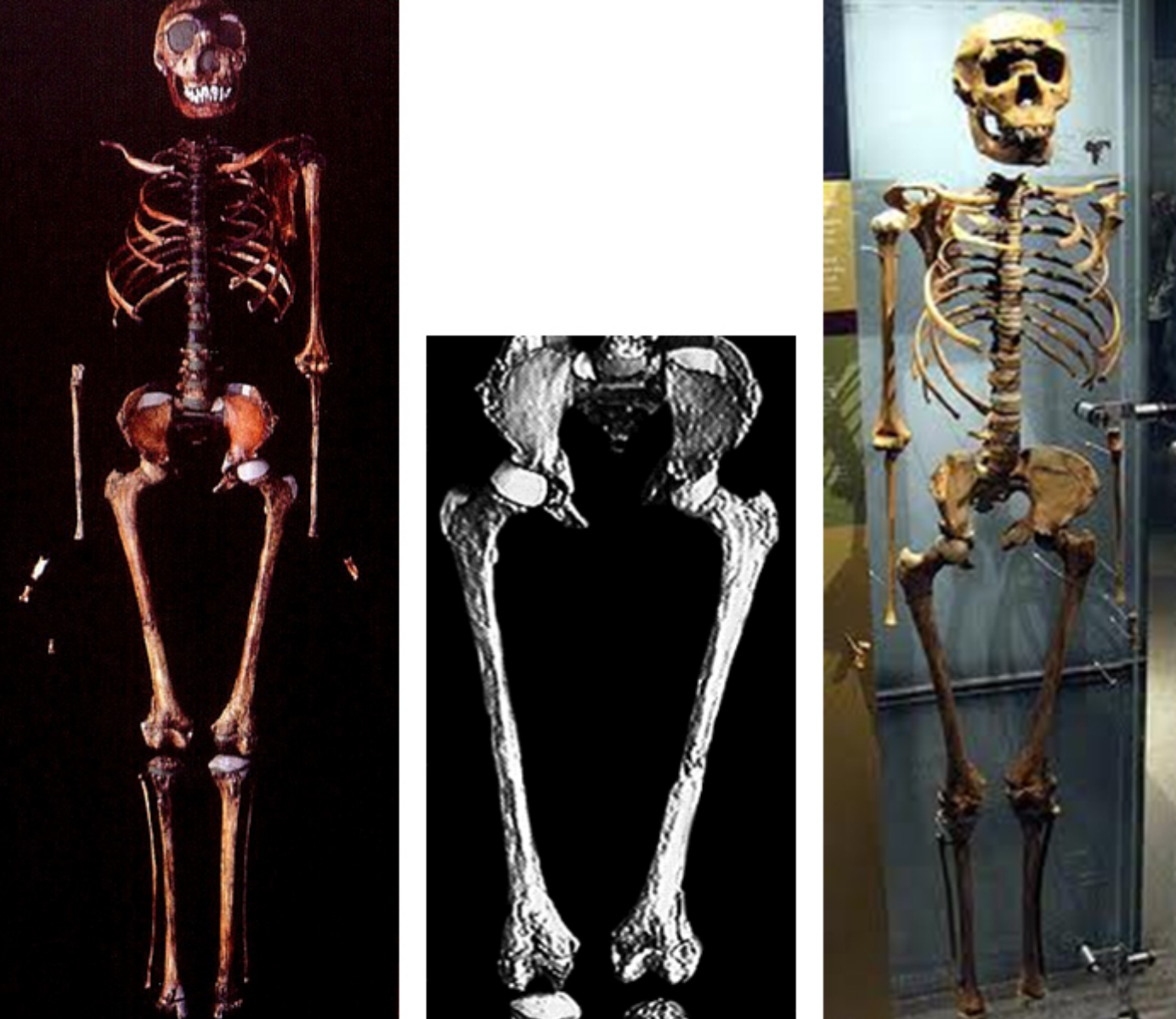 |
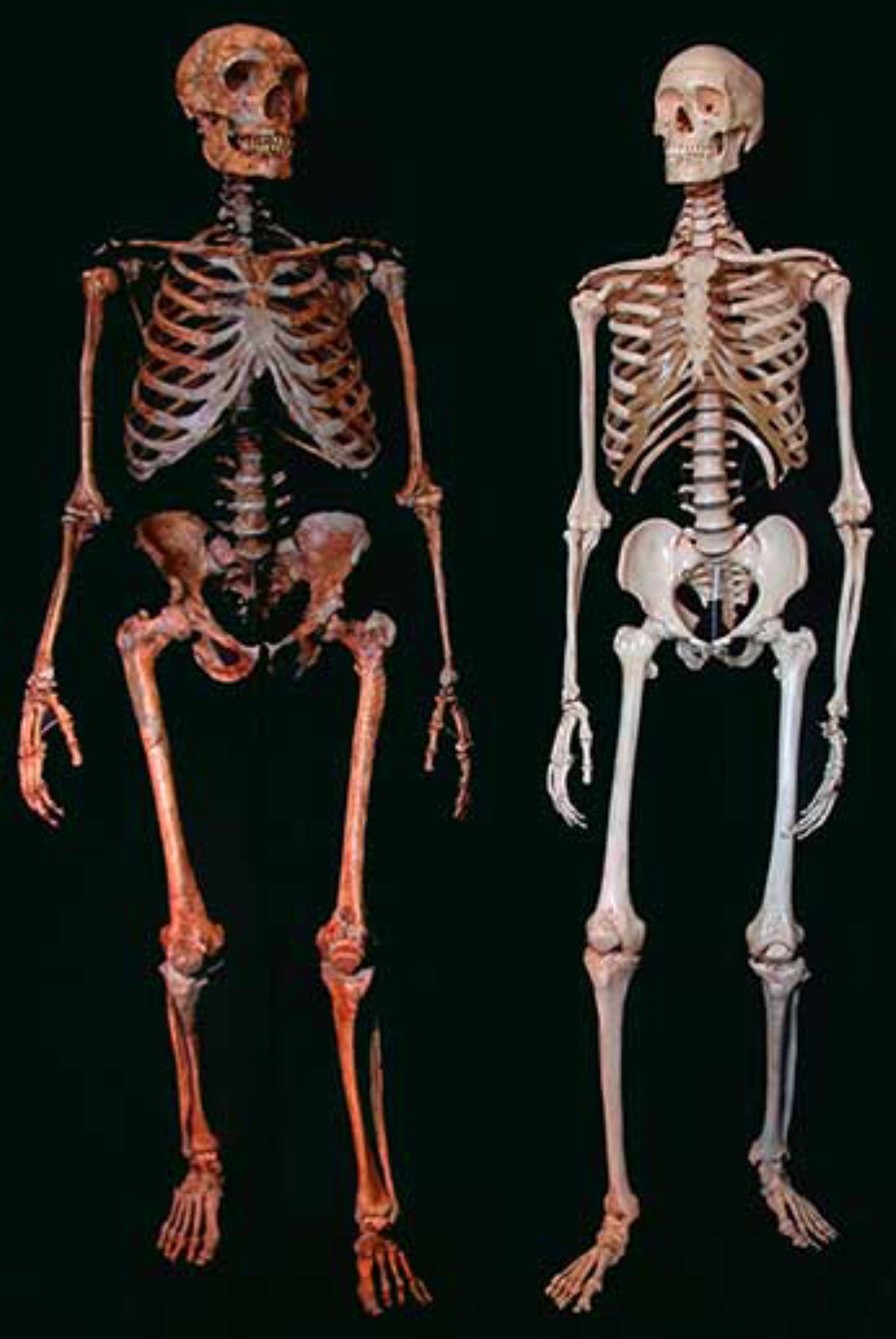 |
Now, let’s talk about legs. Anything distinctive about the legs of pre-humans, or hominoids, or Neanderthals? Anything they have in common? Because if they do, that further indicates they are the same kind of creatures, doesn’t it?
Here is some food for thought. Let’s go back to Lucy, Australopithecus afarensis, at 3.2 mya.
Like all pre-humans, Lucy had a wider pelvis than humans, and of course the flat feet with the forward-placed ankles from the Laetoli tracks.
Building a knee joint in the middle of those two areas of stress — a wider starting point (the hip joint) and a more inward-tilted end point (the flat feet), impacts most on the knee, which has no choice but to slant inward (become knock- kneed) for the legs to function successfully.
Are knocked knees found in any other pre-humans? How about one of the most complete early hominids, the Homo Ergaster known as “The Lake Turkana Boy,” from 1.5 mya.
Here, too, notice the extra width of the pelvis, the sharper inward angle of the thigh bone, and the inward slant of his knees. His feet were not found, but we assume they were as flattened as the others we’ve seen, as per the Laetoli model.
How about a Neanderthal-human comparison?
Again, the Neanderthal’s wider hips and flatter feet force the knees to “knock” inward in order to be able to walk smoothly and comfortably.
Now notice the human’s narrower pelvis, and how it coordinates with the arched foot to allow a straight line with the thigh bones and the shin bones. A completely different leg design, which produces a completely different walking style.
 |
Last but not least, there is one more noticeably knock-kneed individual we have already met:
That’s right, Patty from the Patterson-Gimlin film. Watch her walk in slow motion and pay attention to her right knee motion as she strides. (The left knee’s motion isn’t as easy to follow.)
You will see it bows inward at each step. She’s clearly “knock-kneed,” and I consider it safe to assume Zana was, too, as well as the Iceman.
***
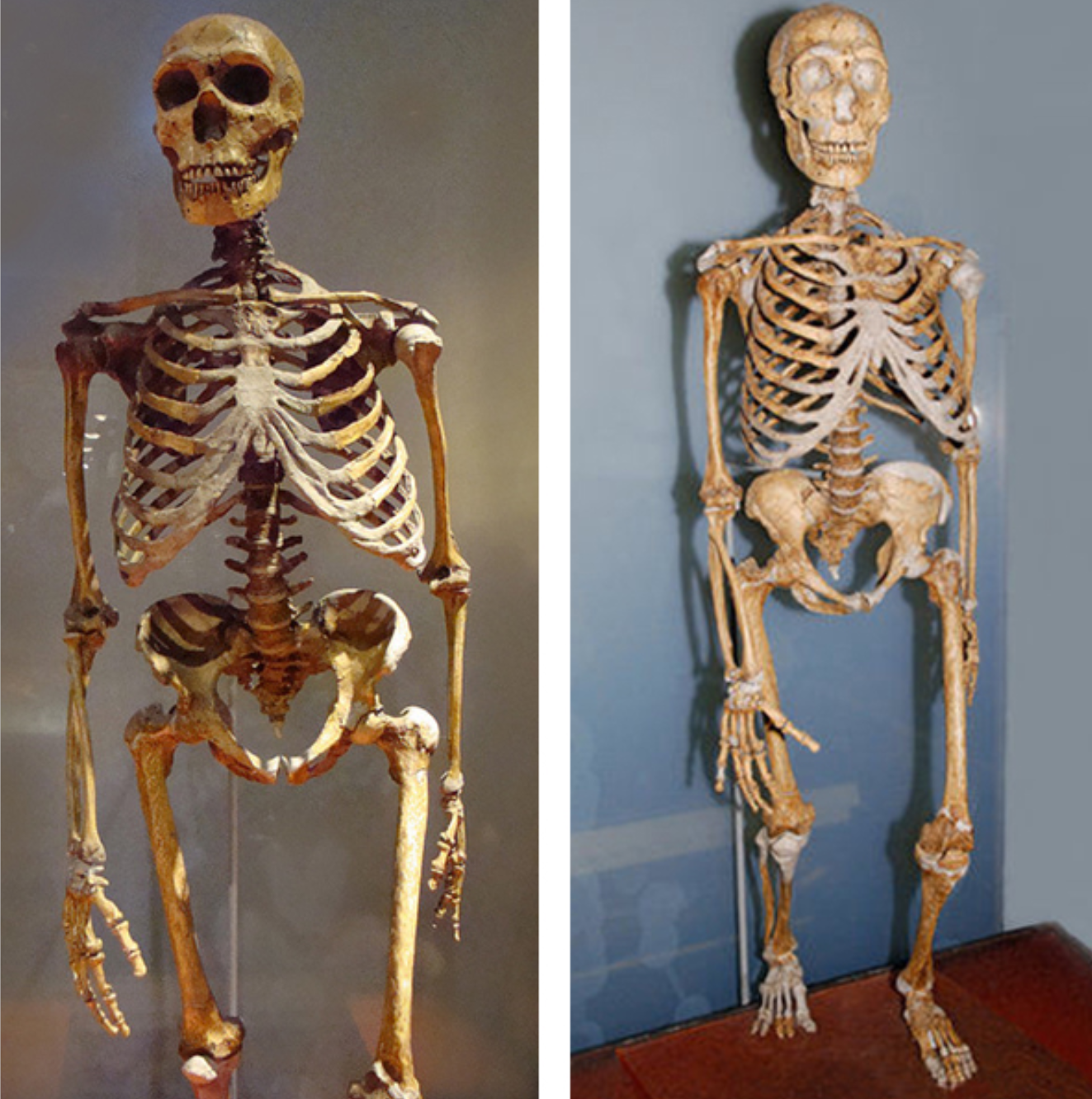 |
While we’re at it, let’s make other comparisons between the Neanderthal and human skeletons above. Neanderthal bones are uniformly more robust than humans. They have Λ shaped rib cages, while humans have ∩ shaped ribs.
Neanderthals have a shorter waist, with only 4 lumbar (lower back) vertebrae compared to the 5 or 6 in humans (“short” or “long” waisted).
Notice, too, the arms, which are crucial. In the Neanderthal reconstruction above, which is said to be from the bodies of several specimens, the arms hang down at a very human-like length.
Contrast that with most hominoid eyewitnesses who mention “extra-long” arms hanging “down around the knees.” With that arm length going back millions of years into the Miocene, we see what has to be considered reliable consistency.
If we can feel safe assuming that Neanderthals might well be the almas/kaptar type hominoid, then I have to wonder if maybe those putting together the model above might have — either deliberately or inadvertently — managed to put young arms, teen arms, on an adult body?
That certainly seems possible, doesn’t it? And if that is what happened, then it stands to reason that the natural arms of Neanderthals could well be a bit longer than is depicted in that model.
In that vein, top right is a Neanderthal skeleton on display at the U.S. Museum of Natural History. Notice its right arm (left side as we view it) is markedly longer than the left arm (right view).
Its right elbow is inches below the bottom of the ribcage, while in the left arm it is even with it. Why? Look carefully at the arm bones.
The left arm is different from the right, with a humerus (upper arm) clearly shorter. Thus, it seems fair to suggest that the two arms might have come from different Neanderthal bodies.
With that said, it is equally important to note that “longer-than-human arms” would not be required to qualify Neanderthals as hominoids.
Maybe some hominoids do, in fact, have arms as long as human arms. But faced with so much evidence to the contrary, I’m not betting on it. I believe Neanderthals are at least one type of the man-size hominoids, and maybe more than one.
The heart of Intervention Theory is its position that humans definitely and beyond any doubt did not evolve on Earth. And, once we became established here (by whomever or by whatever established us), we have since done no more than microevolve into our fully modern form.
What is the evidence in support of this radical theory? It comes in two general categories: the physical indications and genetic indications. Let’s examine both of them in a bit of detail.
Physically, humans supposedly have evolved, or “descended from a common ancestor” that we share with chimps, diverging at 5 to 8 mya.
If that were actually true, we should share many — not all, but quite a few — of the chimp’s most fundamental physical traits. However, when we look under the hood, so to speak, what we find is little in common beyond having 97% of the same genome (our individual gene pools).
For people unfamiliar with genetics, that 3% differential sounds trivial because they don’t realize our respective genomes contain over 3 billion nucleotides (also called base pairs, or bp), so 3% of those would be 90 million bp. If you know what you are doing in a genetics lab, 90 million bp can be a lot of room to roam.
The enormous physical differences between humans and chimps should make it clear to any person with an open mind that someone knew how to make good use of those 90 million bp.
Here is something else to consider in terms of that 97% genome similarity between humans and chimps. Humans have 70% the same DNA as rats, 65% the same as mice, even 25% the same as single-cell, insensate, mindless yeast.
***
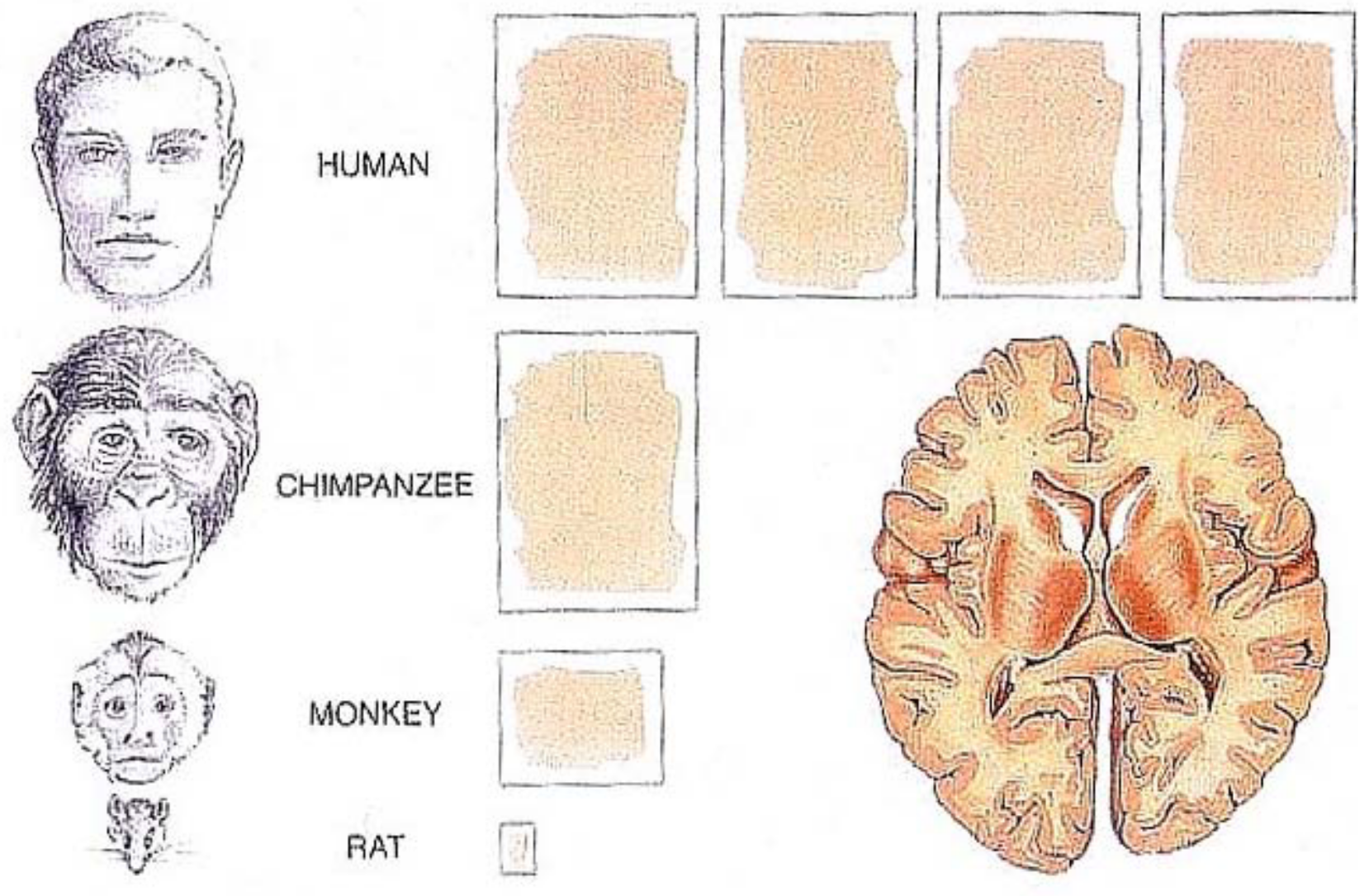 |
 |
There are 12 major — and many more minor — physical differences between humans and the higher primates (HP), like chimps and gorillas. Let’s examine those 12 differences in detail.
1. Locomotion – This is the most obvious. We are bipedal and HP are quadrupedal. This is as different as different gets … enormously so.
2. Skulls, and Brains in Skulls – Here, too, the differences are glaring. HP have brains that are massively reduced in mental capacity compared to humans, and they are also reduced in size.
Top right shows that a cerebral cortex, the surface covering of any brain that handles much of its mental workload, is the size of a postage stamp in rats, a postcard in monkeys, a sheet of typing paper for chimps, and four sheets for humans!
Each case extends well beyond evolutionary transition, it is a transformation, likely more attributable to Terraformers than to evolution.
As for the skulls, all HP (and all pre-humans, for that matter) exhibit the same skull features. 2nd from top is a gorilla (top left), human (top right), and three pre-humans across the bottom row.
All of the HP and the pre-humans exhibit:
- Thick, heavy, protruding brow ridges.
- Foreheads that recede straight back from the heavy brow ridges. No vertical foreheads.
- Large, round eye sockets that will have eyes capable of some degree of night vision.
- Wide nasal openings, which indicate their noses were as broad and flat as all HP today.
- Mouths pushed outwardly away from the face, protruding in a prognathous fashion.
- All HP and pre-humans lack chins, which is a universal trait shared by other primates.
Human skulls are exactly the opposite of each of those traits — exactly 180 degrees opposite!
All humans have greatly reduced brow ridges above greatly reduced eye sockets, which rest below a vertical forehead. Our nasal bones rise up to varying degrees to lift our noses up off of our faces so we can go underwater without the water gushing into our noses. HP can’t do that.
Humans have chins that are the exact opposite of HP’s chin area. Likewise, human mouths are pushed much flatter against the face than HP’s. These differences could not be more dramatic.
3. Bones – We have already learned the major difference in HP bones and human bones. Our bones are much thinner and lighter than typical primate bones. If we did indeed evolve to the condition we are now, where was the adaptive advantage in becoming so much more fragile?
4. Muscles – As we also learned earlier, human muscles are 5 to 10 times weaker than muscles in all HP. Where was the adaptive advantage in becoming more fragile and that much weaker? How could it make us better able to compete?
5. Skin – Light-tone skin is not well adapted to sustained direct sunlight. Even those with dark skin will eventually suffer adverse affects from sustained exposure with no relief from shade or using clothing. All HP have full body hair that protects their skin from unlimited sun exposure.
6. Adipose Tissue – We have 10 times as much fat beneath our skin as HP have. When they are cut severely, their skin will pull itself together and heal as if a doctor had stitched the wound.
If we are seriously cut, the fat under our skin pushes the wound edges away from each other. Without stitches, serious cuts become infected, which throughout history usually meant death. Where was the adaptive advantage in that?
[Most animals on Earth lack adipose tissue, so their cuts heal as easily as higher primates. The exceptions are sea-dwelling mammals, which have varying amounts of adipose fat to provide insulation, which it also provides for humans.]
7. Body Hair Missing/Pattern Reversed – We lack the thick hair pelts of HP, and the patterns of growth on our bodies are the exact opposite of theirs. HP hair grows thickly on their backs and lightly on their fronts. Human males are thicker in front than in back. (Human females have no heavy torso hair.) Why and how would evolution engineer such an extensive switch?
8. Head Hair and Nails – Primates have head hair and nails that grow to a prescribed length for each species and then stop. They never have to trim either. So, why would humans grow hair and nails that must be regularly trimmed? Once again, where is any adaptive advantage in this?
 |
 |
 |
9. Throats – Total rearrangement. Primates can eat and drink while breathing; their windpipe is separated from their esophagus. Humans can’t do it without choking, possibly to death. Once again, this would not seem to hold any adaptive advantage for humans, yet somehow we have it.
[Re: #2, above, noses, the extensive redesign of the human skull permits our nasal openings to submerge and water won’t pour into our throat. HP are not as fortunate. When submerged, the sharp “in-and-downward” angle of their nasal openings lets water gush in to choke them.]
10. Sex – HP have obvious estrus cycles, while humans do not. This is a momentous change on several levels, moving us extremely far from all HP in this vital aspect of the life of any species.
11. Genetic Disorders – Most animal species, including the HP, have relatively few genetic disorders spread throughout their gene pools (from a few dozen to a few hundred). Many of those cause non-fatal afflictions, like albinism, which are passed to offspring without harm.
Severe genetic disorders tend to be weeded out and kept from the gene pool of all the “natural” species (non-domesticated). Humans, however, carry over 4,000 genetic disorders — from mild to severe — and new ones are found regularly!
A couple dozen of those disorders kill all who express them before they can reach puberty to pass them on. So how did those get in our gene pool in the first place? How can they replicate from generation to generation, among every race and creed and culture on the planet?
That answer is well understood. They replicate through “passive” carriers, who first “initiate” the genetic “defect” as a mutation, but in whom it is “silent” and does not harm them while they live. However, if they mate they can pass it on.
Thus, one deadly defect can appear in a random individual, then gradually it can be passed on to enter the gene pool. It happens all the time. But now comes the problem that is hard to explain.
If we humans are a recent species (less than 6 million years old), why do we have so many more terrible mutations than many far older species dating back tens of millions of years (such as rats or opossums or even our friends, the coelacanths)? The mainstream can’t answer this, but I’ll take a stab at it in the next section.
 |
12. Chromosomes – All of the higher primates carry 24 chromosomes from each parent, for a total of 48 in the cells of each individual.
Humans carry 23 from each parent, for a total of 46. So, how could humans “lose” two entire chromosomes and still become vastly “better?”
Before we try to answer that, let’s recall that despite “losing” two full chromosomes from our genome, it has stayed 97% the same as in chimps, and 95% the same as in gorillas. That doesn’t seem mathematically possible, does it?
Well, it isn’t; we didn’t actually “lose” them.
The 2nd and 3rd chromosomes in HP are, in humans, fused together to make chromosome #2. This is an enormous and critical difference, so how could such an oddity have occurred?
Mainstreamers insist Nature is fully capable of creating such a fusion by means of a mutation called a Robertsonian translocation. (These are complex and later will be discussed a bit more.)
They further insist this profoundly mutated HP individual could somehow grow from a single mutated cell with normal cell division, all the way to birth and then onward to reach puberty.
After that, they presume it finds an unaffected mate and reproduces, thereby inserting the new fused chromosome into the HP gene pool as a radical mutation that will gradually disperse.
In the real world of humans and HP, a human-HP sperm-egg combination cannot and never will work. No human has ever mated with a HP to produce viable offspring. And don’t think it has never been tried! Russians in the 1950s and 1960s tried especially hard to find a way to add the “extra strength” of HP to their soldiers.
The truth is that the only way such a massive change could occur among the HP and then be successfully passed on to create a new “line” is if the Robertsonian translocation happened in exactly the same way to different individuals, a male and a female, at virtually the same time.
In addition, those two individuals had to live in the same general area, survive to breeding age, find each other without knowing they carried a game-changing mutation, and then mate to have offspring that could propagate the mutation.
Once offspring are born, they must live long enough to find similarly mutated mates, which at first means mating with mutated siblings to have offspring, while avoiding inbreeding.
In order for the mainstream to maintain their genetic model of human evolution, they must convince us that the particular Robertsonian translocation we have was not fatal to the first individual to initiate it. This is also why they avoid discussing these issues in open forums.
***
Those 12 physical differences between HP and humans are essential aspects of the truth about human origins. They prove human and chimp physiologies have very little in common despite supposedly sharing a common ancestor at 5 to 8 mya, as well as the great bulk of their genomes.
This has been a huge problem for mainstream science since Darwin wrote their evolutionary gospel in 1859. That is a primary reason why they have to work so hard to prop it up, and so readily attack anyone who dares to challenge it.
Nothing supports the Intervention Theory and its Intragalactic Terraformers quite as much as the fascinating genetic differences between our human DNA, and chimp and gorilla DNA; and, since recent recovery, Neanderthal DNA; and, at some point in the future, hominoid DNA.
As we explained a few pages ago, the second chromosome in humans is a fusion of the 2nd and 3rd chromosomes in higher primates (HP).
The mainstream claims it as caused by a rare mutation called a Robertsonian translocation, which can combine chromosomes end-to-end, telomeres-to-telomeres, to somehow make them function well in a radically new configuration.
To combine two chromosomes so they can keep working is such an incredibly complex series of events, if it were not for mainstreamers having a desperate need for that event to be considered plausible, they would laugh it out of existence.
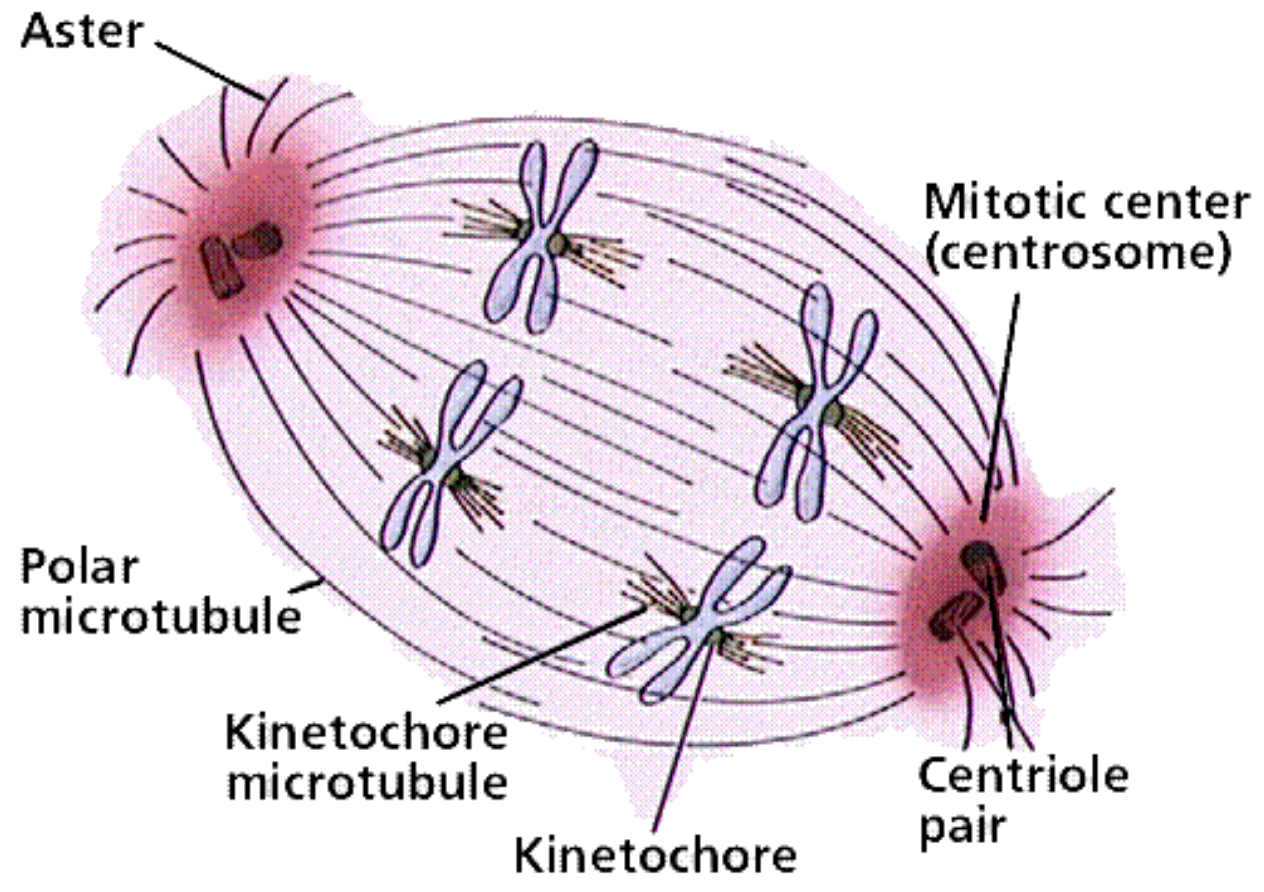 |
Let’s try to follow their logic. Since all HP have 48 chromosomes (24 from each parent) it seems safe to assume any “common ancestor” (CA) of chimps and humans had 48 chromosomes.
Let’s assume two CAs have sex, and somehow in that process, the female’s egg has undergone a Robertsonian translocation mutation and its 2nd and 3rd chromosomes have fused into one.
When that mutated egg meets any normal, 24 chromosome sperm, it will not form a fertilized zygote … or if it does, soon after it will expire. Why? Because to replicate into more cells, each chromosome must line up with its pair from the other parent before being duplicated and pulled apart by fibers to opposite ends of the cell.
Then, the one cell splits into two. This process is called mitosis, and every one of our trillions of cells are copied in this manner, one after the other after the other, from the original one cell.
Now, if we consider a human-chimp cross, the 2nd human chromosome must line up with two chimp chromosomes. However, as the contents of the cell first duplicate and then pull apart, the intricate “dance” between them will soon end.
Why? One copy of the human 2nd chromosome and one copy each of the chimp 2nd and 3rd must somehow safely wind up at one end of the cell, while the other three copies need to be pulled to the other end before the cell can divide.
In that process the fibers become confused, and the resulting cells try to keep on replicating, but that chaos continues until the blastula expires.
So, there is no way that one-chromosome-short zygote will somehow become a viable fetus.
 |
In addition to the above, now let’s consider the problems found in telomeres and centromeres.
Telomeres are the “caps” found at the ends of chromosomes that gradually reduce after each cell division. Think of them as a long string of “beads” on a necklace, and after each division of each body’s trillions of cells, a bead is lost.
When all of the telomeres have dropped off, the chromosomes stop replicating and the organism they support will die from advanced “old age.” Nothing can stop the slow loss of those beads.
Centromeres are segments of DNA usually located near a chromosome’s middle, and they are critical to successful cell division, which is the continual process of life that has to happen correctly, each time, every time, or things can go very, very haywire within the organism.
Now, with that in mind, let’s try to imagine what would happen if a pair of chromosomes fuse in the way the two primate chromosomes fused to create the “missing” one in humans.
The fusion puts the two central telomeres (blue) into the middle where the centromeres should be, and the new chromosome has a pair of (red) centromeres when it should only have one, and that one should be where the telomeres are.
This is a serious problem because telomeres perform a “stopping” function that is entirely inappropriate in the middle of a chromosome that is supposed to be fully functional. Uh-oh!
Even worse, the centromeres are only useful in cell division, so when that occurs there will be not one, but two places where it is happening, which will soon lead to a badly tangled mess.
Clearly, mainstreamers need multiple miracles to make this scenario plausible … and guess what? The exact array of miracles required has been found within human chromosome #2!
Traces of two HP telomeres are found in human chromosome #2, between bases #114,455,823 and #114,455,838. Those 15 are deactivated in some way that doesn’t stop the chromosome’s normal functioning. They have been neutered!
With the fused chromosomes, only the middle two telomeres are “deactivated.” The one at the top end and the bottom one at the other end are not altered, so their crucial role in cell division (dropping “beads”) will continue unhindered. How amazing is that? How … coincidental?
As for leaving two centromeres where only one can function, guess what? One of those seems also to have been deactivated, so that normal cell division can proceed successfully! Wow!
The sequential precision of this incredible, one-in-trillions fusion forces us to describe it as yet another of the many miracles the mainstream always seems to be blessed with. Incredible!
 |
 |
While the fusion “miracles” torture credibility for anyone except mainstreamers, believe it or not we find several more in other chromosomes in the human genome! These are inversions.
An inversion can occur when a segment of any chromosome is sliced into, top and bottom, and then pulled out, inverted, and put back into its original place, but with a “flipped” orientation.
According to textbooks, inversions are caused by “ionizing radiation” that causes the genetic bonds of chromosome’s to “temporarily” break loose, during which inversion occurs, followed by a reinsertion. These are rare, but verifiable.
Also consider that any two chromosomes might accidentally become “entangled,” and the result is the brief tearing loose of one segment that then inverts and moves back into its place.
The beauty of this is that every inversion is unique, and if passed on creates a landmark DNA signpost which cannot be reversed back to normal in future generations that carry it. It also works to disprove Darwin, as we shall see.
In theory, while an inversion changes the order of the alleles that comprise chromosomes and genes, the overall makeup of both will remain unharmed as long as every gene temporarily segregated from the chromosome is retained in the process of inversion and reinsertion.
Now, brace yourself for this: The genome of every human carries nine of those “miraculous” inversion/insertions that are not found in any of the corresponding chimp chromosomes! They are located in these human chromosomes: 1, 4, 5, 9, 12, 15, 16, 17, and 18!
According to Darwinian evolutionary gospel, this means that at some point after the proto-humans and chimps split from their supposed common ancestor, the first of the 9 inversions occurred, eventually to be followed by 8 more.
For example’s sake, let’s assume that the first occurred in chromosome #1, at 5 mya. One of the new proto-humans carrying those fused chromosomes gives birth to a child with an inversion/insertion not carried by chimps in chromosome #1. At 5 mya. Simple enough.
Now that child must run the gamut of infant and child mortalities to reach maturity. It does, and then finds a mate. Unlike the chromosome fusion case, which won’t allow offspring with a partner having a loose chromosome, inversions can be passed on with a partner who lacks it.
In each pairing, the offspring will have a one-in-two chance of inheriting the new inversion. The same will hold true for their offspring if they carry it, so their odds of passing it to any one of their children would be 50% - 50%.
Now imagine some astronomical odds. What the above means is that somehow the individual with the insertion in chromosome #1 produced a line of offspring that passed it down to every descendant member of its species to become a part of nearly 7 billion humans alive today!
Since today we all have an identical insertion in our #1 chromosome, it means the insertion had to start at some point with a mutation in one of us who somehow bequeathed it to the rest of us.
This mutation, whether it did something good for the individual who had it, or bad, or nothing at all, would, according to Darwinists, create an aberration that mushroomed out into humanity like a nuclear bomb. Now hold that thought.
For as unlikely as all that is, guess what? The mushroom cloud inversion improbability had to occur in exactly that way for every one of the eight more times it occurred! That’s right, it is, in fact, massive improbability to a power of 9!
In a few million years, on 9 separate occasions, proto-humans were born with a new and quite distinct inversion mutation that would then be passed on to every human alive on Earth today. So the odds of that occurring are enormously more than long, they are beyond imagining!
With all that said, here is the kicker, the thing that will lay you low if you’re not ready for it: Each one had to happen in a sequence! If they all occurred together, it wouldn’t be evolution.
Let’s get clear on what the mainstream insists had to happen. Inversion #1 occurred and the genetic lines of all other proto-humans had to die out. Only its progeny would live to pass the inversion along to subsequent generations.
Next, let’s say that at 4.5 mya the inversion in chromosome #4 occurred, and, of course, that one has to occur in one of the progeny in whom the first inversion occurred or it wouldn’t carry on down through history with the other eight.
Now only its progeny, carrying the inversions in chromosome #1 and #4, can move forward into the future. All other proto-human lines at 4.5 mya have to die out in one way or another.
If the mainstream is right, and the 9 randomly generated inversions occurred in a Darwinian sequence of gradually accumulating mutations, it couldn’t happen any other way. However, is there another way? One that makes more sense?
Of course there is!
What if Intragalactic Terraformers decided, for whatever reason, to insert a “superior” creature here on Earth, a species that would be what we call “human”? Forget microevolution, that’s too slow. They would want to make it happen now!
They have two choices: bring or build. If they decide to bring what they want, they will locate a suitable species — one or more — living on a similar planet. This assures them it can survive here, but to make certain their efforts won’t be wasted, they might bring in several samples to find which one, or ones, manage to adjust best.
If this path is the one they followed, it could explain the wide range of physical variations in the human races. However, from the genetic standpoint, only slight differences separate one race from another. Despite how hokey it seems, under our skin we really are the same species.
If the Terraformers chose to build a whole new species from scratch, it would not be beyond their reach. If they had the technological ability to transform Earth for 4.5 billion years, then it is safe to assume they had the ability to handle genetic engineering at any level of complexity.
So, whether it was a case of “bring” or “build,” the Terraformers could easily have handled it. Which did they choose? And why? Why us? Why humans? What could they possibly have been thinking when that decision was made?
Unfortunately, possibilities and permutations for that answer require an eBook of their own, which I will have to write at another time. For now, no succinct answer covers all the bases.
***
This eBook has expressed many reasons for my firm belief that humans did not evolve on Earth. I am convinced we resulted from sophisticated genetic engineering, the kind we ourselves are only beginning to understand how to carry out.
As proof of my claim, I can point to the fusion in our second chromosome, and the inversions in nine of our other chromosomes. But we also have a few more genetic anomalies that make it clear we don’t descend directly from primates. If we consider the entire genomes of humans, and of chimps and gorillas, we find a startling anomaly. [I’d prefer to include Neanderthals in this, but for now the information is unavailable. However we expect it eventually, and when it is available, I’m confident it will show they were much more like higher primates than humans.]
Chimp and human genomes share 97% to 98% of the same nucleotides (base pairs). It’s a small fraction making a big difference. Both genomes have about 3.0 billion base pairs, which makes the difference total 60 to 90 million base pairs.
Even the low estimate of 60 million differences provides a lot of room for genetic innovation if you know how to carry it out. And most, if not all, manipulation would have to be done on the female eggs because sperm are vastly smaller.
It is also important to understand that humans have 20,000 to 30,000 genes in their genome, and chimps and Neanderthals are assumed, but not yet proven, to have approximately the same.
Within our 20,000 to 30,000 genes are 223 that have no analogues in any other animal species, although 113 of the 223 can be found among bacteria. Thus, mainstream science concludes that all 223 of the “unique” human genes have somehow been put into our genome by “lateral transfer,” which means infection by bacteria.
Well, okay … maybe. But what about the 110 that aren’t found anywhere? Not in animals or plants or in pond scum? Where did they come from? How did they get into our genomes? In bacteria that have not yet been discovered?
I believe those unique genes, and the fusion of two primate chromosomes, and the inversions in nine of our chromosomes, have all come to us through Terraformers who, for reasons and by methods we may never understand, decided to create us (or to bring us) to live on Earth and grant us dominion over every other thing on it.
***
 |
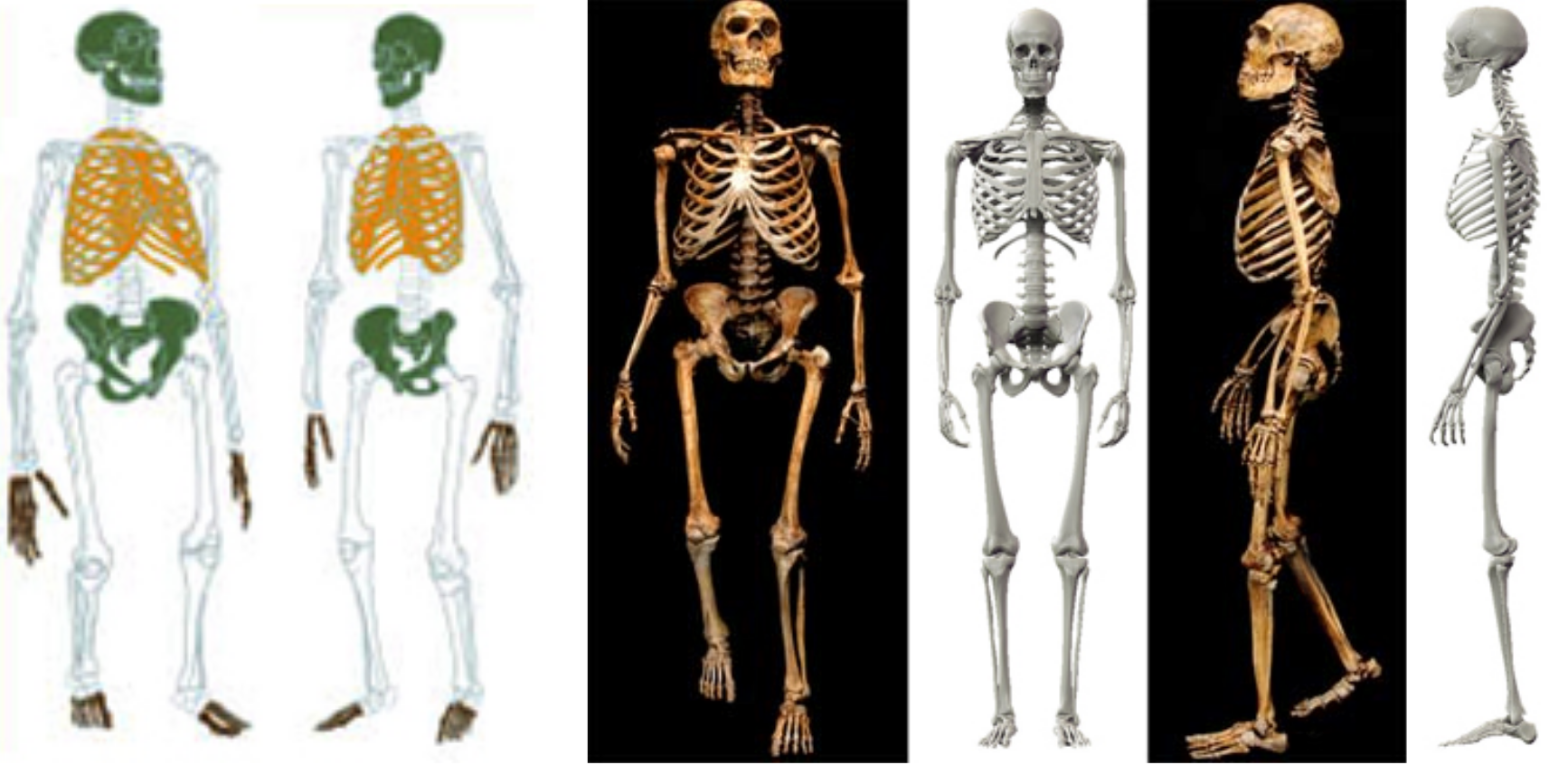 |
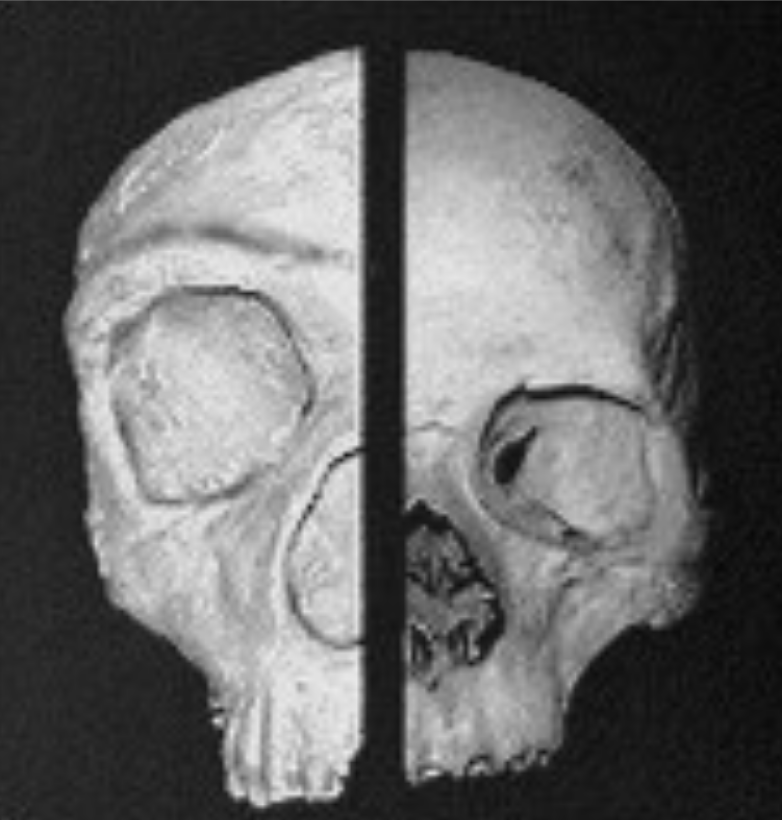 |
What we have discussed above is only the tip of the genetic iceberg that separates humans from chimps and Neanderthals. For example, here is an easy question: How many chromosomes did Neanderthals have? Researchers have most of their genome sequenced, but they never tell us how many chromosomes they had. Why?
Because mainstreamers don’t want to deal with the probable answer — 48. If it were 46, it seems safe to assume we would have heard it shouted from rooftops by now. If it were 46, that would be the same as humans, which would strongly support macroevolutionary theory for humans.
Inasmuch as we are not hearing about it, I have to suspect the answer is 48, and if it is 48, then Neanderthals were vastly more like primates than humans. Furthermore, Zana was likely the Neanderthal that I and others believe she was.
Notice how easily a Neanderthal skull (top right) fits into a primate profile. This would no doubt be true for all of the so-called pre-humans that the mainstream insists are our direct ancestors.
Also keep in mind how strikingly different are the physical variations between Neanderthals and humans. You just can’t make this stuff up.
This single illustration (bottom right} might make it as clear as it needs to be made. Neanderthal on the left, human on the right. Need I say more?
***
Adding all this together, we make a strong case for humans being the result of “tampering” with our genome at some point in our collective past.
Naturally, no one from mainstream science will give Intervention Theory an iota of credibility, but in many ways their knee-jerk rejection of it provides one of its strongest bases of support.
If this eBook teaches readers anything, it should be that whatever the mainstream says about the sensitive topic of “origins” should always, and invariably, be taken with a large grain of … no, make that a wide-brimmed bucket of … salt.
Scientists are, at bottom, tentative. They have to be or they can’t survive, or rise, in a system that requires undoubted loyalty to dogma. This does not permit them to think or act independently. It forces them to always put dogma before facts.
Given the choice of humans being created by genetic engineering, or various races brought here from other planets much like Earth, I lean toward us being engineered as a stock species.
MtDNA studies suggest the base species was blacks in Southern Africa. Then the other races could have developed from extended intervals of isolation in various areas around the globe.
[During those millennia gradual microevolution (a la Darwin’s finches) might well account for the slight physical changes in different races.]
It also remains possible that our races resulted from periodic rounds of genetic “tinkering” by Terraformers to “modify” the original “model,” for whatever reasons they might have had.
We might also have resulted from hybridizing between similar genetic species, including the Terraformers. (For those wanting to know more about this controversial idea, I can recommend The Earth Chronicles, by Zecharia Sitchin.)
However the human races came to be on Earth, as that was unfolding, the bipedal apes that had dominated the planet until then continued living their lives in the geographic areas dominated by forests, woodlands, and jungles — well removed from the open areas the new beings preferred.
Then, as now, occasional encounters between the two would occur. The full accounting of hominoid history, like the full accounting of human origins, demands an eBook separate from this generalized accounting of both.
In the end, for as convincing as is the evidence for hominoid reality provided by the Patterson-Gimlin film and the Minnesota Iceman, some of the very best — if not the best — evidence for the Intervention Theory remains … Zana.
***
Clearly, something astounding happened when men of Tkhina had sex with Zana and produced normal human offspring. Typical hybridization did not occur. The rules went out the window.
She was a bipedal higher primate from deep in the Miocene, yet her offspring with the men of Tkhina — while exhibiting traces of her outward characteristics — were overwhelmingly human. How is that possible? How can it be explained?
If, as I’m convinced, humans are the product of genetic engineering by Terraformers, then it is likely that the Terraformers created, or placed, bipedal Miocene apes on Earth at 23 mya. If we assume that happened, the Terraformers knew the hominoid/Neanderthal genome inside out.
When creating humans, maybe they took a few “shortcuts.” For anybody that can use highly sophisticated techniques to manipulate DNA, a “designer baby” would be easily within reach.
Maybe the Terraformers saw Neanderthals as well along toward what they wanted their new “upgraded” primate to be. Maybe for them it was like “souping up” a car, turning a stodgy street sedan into a far more nimble “hot rod.”
Whether a valid image or not, the point is that Neanderthals could have provided a functional template to construct the new species. Smaller, weaker, more fragile, not covered in protective hair, and having several other physical short-comings — but vastly smarter and more lethal.
[We have already discussed how different we humans are from primates, so the question is still open as to how and why that came to be.]
Getting back to Zana, if she was indeed a sex slave during her 40 years in Tkhina, she might have had sex with local men and boys several thousand times, yet she had only 8 pregnancies. Furthermore, in each case, her offspring were far more human than hominoid/Neanderthal.
An oddity of those pregnancies is that Khwit, Zana’s youngest son, was born in the middle-1880s, when she had lived in Tkhina for about 35 years, and about 5 years before she died of apparently natural causes. We don’t know her age upon capture, but she seemed fully adult.
A trait that distinguishes primates from humans is that among females, only the oldest primate females tend to stop having estrus cycles, and usually shortly before they die. As best we can determine, Zana seemed to follow that pattern.
In some ways, my assumptions are what I often accuse the mainstream of doing, which is to get results they want by working backward from the starting point of a predetermined answer.
My known answer is that Zana was clearly not a human female, she was a hominoid, and very likely a living Neanderthal. Yet she mated with humans to create dominantly human offspring.
This is not how normal hybridization works, so perhaps something was “inserted into,” or was “removed from” the genomes of each species to allow their hybridization to occur as it did.
How? Why? That is really anyone’s guess at this point. However, it’s worth noting that only very recently, in 2010, did genetic researchers discover that the genomes of all non-Africans contain up to 4% of Neanderthal DNA.
This very strongly suggests that humans have been able to mate with Neanderthals since we first appeared on Earth … OR … and this is a larger step to propose … or Neanderthal DNA was manually inserted into our genomes by the process of genetic engineering that created us.
That suggestion leads to the idea that we are so radically different from any other primate on Earth because, fundamentally, in our genetic core, we are not of the Earth! We could very easily be dominantly from somewhere else.
This brings us back to the question of whether life as we know it is ubiquitous throughout the universe, or at least within our galaxy. I believe it is because the genetic structure of every form diligently conserves itself, and multiplies itself.
If I’m correct, then it isn’t too much to presume that several genetic aspects of Terraformers — if not many aspects — could be exactly the same as ours. We could well be made in their image!
As it happens, this claim was also made in the Bible: Let us make man in our image, after our likeness. Exchange the word “man” and put in “Adamu,” and it’s almost a direct lift from a Sumerian text written 2,000 years before it!
So, is it possible? Can we humans be “aliens” living on Earth, created or brought by entities we call Terraformers, with a genetic package very similar to Earth-based primates because all species throughout the galaxy and universe are created from the same genetic code of life?
Let’s work back from another predetermined answer: Humans do exist on planet earth. We seem to have existed as we are now for 200,000 years, if Early Modern Humans can be counted as us, which is not necessarily how it happened.
If the Cro-Magnons are the actual beginning of modern humans, then we’re only about 60,000 years old, which contradicts the mitochondrial DNA studies that conclude it is 200,000 years. However, either date demolishes the “common ancestor” theory that dates us from 5 to 8 mya.
Whenever we appeared, we have looked and acted like nothing else on the planet. We have the genetics of this planet’s primates, but that can be very misleading in terms of functioning.
This planet’s primates, including us, could have the basic genetic package of many other species in the galaxy. Just as life is here on Earth, our genetic code could exist absolutely everywhere!
After all, 95% of human DNA is, at this time, considered “junk” by scientists because it does not code for proteins the way “working” DNA does. Nonetheless, it remains extremely highly conserved, which means our bodies continue to reproduce it, so it can’t possibly be actual junk.
Because of science’s fundamental and profound ignorance of the reality we live in, of what is in our bodies and how it works, of where we have come from and where we are going, they will continue doing what they have always done.
Rather than investigate, they will castigate, and that will be their tactic until they are confronted by evidence so overwhelming, it will end their game of denial, and their jig will finally be up.
Intervention Theory requires certain things to be true for it to be acceptable as the most likely explanation for how all of life, and particularly how humanity, has come to reside on Earth.
Intervention and evolution cannot coexist — they are mutually exclusive. Unfortunately, Intervention incidentally supports Creationism, which I regret because, while religious dogma was understandably a community staple 5,000 years ago, and 2,000 years ago, and 200 years ago, it is no longer rational in real-world terms.
If religion could be separated from the search for actual truth, science might be more at ease with confessing their profound ignorance about so many of life’s verities. But because so many religions breathe down their necks, they don’t have leeway to admit mistakes or to be wrong.
This doesn’t mean science is without blame in the matter. Compounding the constant pressure applied by disgruntled religious zealots is the hubris and arrogance that always comes with the rank and power of positions of authority.
If the truth about life on Earth is what we seek, then I believe Intervention Theory best fills the bill. And I believe that truth is best expressed in the analysis of human history we’ve carried out in this eBook, especially in our relationship, the degree of our relationship, with Neanderthals.
If, as I have suggested, Terraformers used the Neanderthal genome as a template to create humans, then much of their genetic package — the genetic package of all higher primates on Earth — would be found in the new humans.
If we assume, as I do, that Neanderthals carry the 48 chromosomes of higher primates, then their 2nd and 3rd ones would have been precisely fused to make humans what the Terraformers wanted them to be. How precise was that?
During the Robertsonian translocation fusion, in another of the miracles the mainstream keeps pulling out of their hats, no genes from either chromosome seemed to be lost or inactivated!
However, as seen above, an insertion occurred with several of the 223 unique genes not found in any other animal species. Scientists claim it is not clear if those genes “provide a selective advantage,” which means they can’t figure out how to make them fit into evolutionary theory.
Well, let’s wish them good luck with that, while we note that taken together those inserted genes contain about 150,000 base pairs that do not, in any apparent way, damage the functions of the newly fused chromosome. Another miracle!
This suggests that while a perfect reproductive “fit” no longer existed between the species with 48 chromosomes and the species with 46, none of the actual chromosomal material is missing.
Everything needed to create offspring between the two species is still available and functional, but the components have been shifted around and reconfigured into a novel arrangement.
Thus, while offspring would not be the likely outcome with any sexual union, it would not be impossible. All the ingredients are there, and if put together just right, offspring could result.
So far as is known, humans and chimps have never “mated” successfully, but such “trysts” do not seem like “high number” occurrences. In a genetics lab … that is a different matter.
I can’t say with certainty that it never happened in a lab, in vitro or otherwise. But we know the Russians (and possibly Nazis) tried it, and tried seriously, with no reported results of success.
On the other hand, Zana is not the only almasty alleged to have produced offspring. Other cases are on record and are reasonably well regarded, though of course not like her unparalleled case.
It is worth noting that only cases involving the alma/kaptar hominoid types are said to produce offspring with humans. Bigfoot/sasquatch types have not, nor have the snowman/yeti or pygmy types, although they seem the most likely to do so because they appear the most human-like.
So, what was it about Zana and the other almas like her who are supposed to have given birth to humans that express only slight traces of their mother’s genetic heritage as a bipedal primate?
I have to assume it is because her species was the prototype the Terraformers used to transfer the Neanderthals’ very small part (up to 4%) of humanity. And as we saw earlier, much of that 4% helps humans to fight Earthly pathogens.
This does not account for Africans who lack Neanderthal genes, and since the first of us did appear in Southern Africa, it might be safe to assume their group had a different inoculation, perhaps different genes from a different bipedal primate yet to be acknowledged or discovered.
Regardless of how the inoculation process was handled, it seems the original “stock species” from which all modern humans spring was not from Earth, and therefore not prepared to cope with the various diseases, bacteria, and viruses the first of the new humans would encounter.
Throughout human history, the worst thing to happen to any native population was when they were invaded by other humans with pathogens they had never encountered before. Millions — perhaps billions — have perished from it (think Columbus in the New World). So, let’s assume the Terraformers knew what they were doing.
***
I will end this eBook with a remarkable letter sent to me by email on February 12, 1999, from a man who, to protect himself from vindictive peers, called himself “DNA Deep Throat.”
He is a highly skilled geneticist whom I have since come to know by name, though he still must stay anonymous because his mainstream geneticist peers remain as vindictive as ever.
This man was instrumental in the early days of my dealings with the now well-known Starchild Skull, but in 1999, I was just getting started and in much need of help to figure out what it was.
[For more understanding of the Starchild Skull, a book about it, The Starchild Skull, is available at www.StarchildProject.com, and also through www.LloydPye.com, as is an eBook about it.]
An early theory about the Starchild was that it might be a human-alien hybrid, and a question that kept being asked was, “How can a human and an alien hybridize? Won’t they be too far apart genetically? How could that happen?”
As we have seen, that question is essentially the same one we have to answer in regard to Zana.
With both Zana and the Starchild in mind, read the words sent to me in 1999, and you will have the same degree of insight they imparted to me.
Dear Mr. Pye:
I agree with your conclusions and will give you a few hints, if you wish, (speaking) as a “DNA Deep Throat.” First, look up the huge discontinuities between humans and the various apes for: (1) Whole mitochondrial DNA; (2) genes for the Rh Factor; (3) and human Y chromosomes, among others.
Regarding #3, I refer you to K.D. Smith’s 1987 study titled “Repeated DNA sequences of the human Y chromosome.” It says “Most human Y chromosome sequences thus far examined do not have homologues [same relative position or structure] on the Y chromosomes of other primates.”
Human female X chromosomes do look somewhat apelike, but not the male’s Y. This means that if humans are a crossbred species, the cross had to be between a female ape-like creature (i.e, “creature of Earth”) and a male being from elsewhere.
What the evolutionists do is find certain genes which look very similar between man and ape, then they make a “tree of descent” while ignoring those huge impassable abysses of difference elsewhere.
Also, by certain methods of DNA dating, one can tell that numerous genes have been recently added to the human genome.
If workers in my field were to say such things openly, we would be ostracized and forced to live in a tent. Any work along these lines would be rejected without any form of appeal. So what can we do?
Sincerely,
DNA Deep Throat
Let’s leave it there, with DNA Deep Throat’s parting question: What can we do when we are up against a monolith like mainstream science, which won’t forsake its cherished dogmas until those dogmas are crammed down their throats?
The End
 |
Lloyd Pye was born in 1946, and raised in Amite, Louisiana, USA. He attended Tulane University in New Orleans, graduating in 1968 with a degree in Psychology. He then joined the Army and became a background investigator.
In his 20s, he worked at several jobs before deciding to write. He sold his first novel at 30, then spent much of the 1980s screenwriting in Hollywood, achieving only moderate success.
In his 30s and 40s, he developed an alternate theory of human origins based on the notion that bipedal hair-covered hominoids (bigfoot, yeti, etc.) were living descendants of ancient pre-humans, and modern humans don’t appear on the flowchart of natural life on earth until Early Modern Humans at 200,000 years ago.
 |
In late 1997, Lloyd published Everything You Know Is Wrong, a nonfiction book detailing his unconventional theories about human origins.
It was a bestseller in Alternative Knowledge, and is priced at $19.95 at www.LloydPye.com. [Amazon.com sells only used copies of it, and the prices they charge for it are outrageous.]
5 out of 5 stars THANK YOU LLOYD PYE FOR WRITING THIS BOOK
By Anne Teachworth
This book makes more sense than anything I've read since “Chariots of the Gods” came out many years ago. I have gotten more answers to Biblical mysteries, Garden of Eden stories, creation tales, Sumerian history, human evolution, and countless other wonderings that Lloyd Pye has carefully explained in language even a novice to this field can comprehend.
I guarantee you will be enlightened and intrigued by all the information he presents about our human-gods genetic ancestry. I stayed up all night reading this book the first time I opened it. I have bought eight copies to give my friends for Christmas, so they all can move forward in life more fully informed and with their minds opened.
 |
Lloyd has also published The Starchild Skull, about his first 8 years as research coordinator for The Starchild Project. A 280-page paper-back, find it at www.StarchildProject.com.
5 out of 5 stars As revolutionary as Darwin's "On The Origin of Species"
By Ralph Hollister
Imagine yourself in Europe 1,000 years ago and you found photos of the Earth with the Moon in the background taken from the Space Shuttle, and you had enough wisdom to understand what they were. Now imagine trying to get anyone to accept what we all know now as a basic truth in our reality.
Since those times much has changed, but a great many things have stayed the same. This book chronicles Lloyd Pye's decade of struggles to solve the mystery of this weird skull that he is convinced is of an ET.
If nothing else, this story is a lesson in how the scientific community circles its wagons when faced with something that could contradict any of its accepted dogmas. But it is much more than that. I didn't think there was anything to UFOs prior to reading this book, but I must admit I am now, like Lloyd Pye, very anxious to find out all there is to know about this unique skull.
 |
Lloyd has also published two popular novels, both available through his websites. At right is the cover of Mismatch, a Cold War spy thriller in the vein of The Hunt For Red October. It is a unique mix of white-knuckle action and inside communications info that Lloyd had access to.
5.0 out of 5 stars One of the best hacker and phone phreaking fictions available.
By A Customer
In Mismatch, Lloyd Pye adeptly tackles a story painfully abused in recent fictions. That is the technical suspense involved with computer hacking and phone phreaking. Movies from 'War Games' to 'The Net' and 'Hackers' present an inaccurate portrait of how hacking occurs and how it can affect the general population.
Mismatch gives a realistic feel to its coverage of technology, and the persons involved in it. With acknowledgements to Naval Intelligence and John “Cap'n Crunch” Draper, King of the Phone Phreaks, you know Pye did his homework. The story is a Clancy-like suspense thriller involving the internet’s early days.
There are a large number of characters, with the most interesting being the lone hacker who intends to shut down the U.S. military network (a la Robert Morris) by blending hacking and phone-phreaking. Naturally, there is a dogged telephone company tracker hunting him down.
A great success of this book is Pye's ability to depict his characters believably. Many others have failed. If you are interested in a fast-paced techno thriller, and not some pseudo intellectual portrayal of hacking as an arcade game, this book is for you.
 |
Lloyd’s other popular novel is A Darker Shade of Red, based on his actual experiences playing football in college. It is an unflinching look at college football from the bottom looking up, when coaches had to force weak players to quit. Critics call it a book that “fills your heart, then rips it out.” Available at www.LloydPye.com.
5 out of 5 stars I lived through the basis of this story with the author in college.
By Ygnacio “Nat” Toulon #72."
A Darker Shade Of Red
For anyone who loves football, this book’s postscript alone is worth the price. It has insights into the tough mentality of coaches that are right on target.
I played high school football against Lloyd Pye in the Louisiana State semi-finals in 1962 (we won) and again in 1963 (they won), then joined him as a freshman player at Tulane University in the fall of 1964.
Although the characters in his novel are a combination of many personalities we knew and played with, as are the coaches and the story’s main events, it is definitely a riveting tale with incredibly accurate insights into the circumstances we endured.
I can honestly say that the emotions he has presented in this novel are raw, gripping, and exceptionally truthful to our mutual experiences at Tulane. I recommend it.
 |
Lloyd Pye has written one prior eBook called Starchild Skull Essentials, available only at www.starchildproject.com. It delivers all of the essential information about a 900-year-old skull found in Mexico in 1930, and now proven to have both human and nonhuman DNA. Anyone looking for proof of alien intervention in the lives of humans would be wise to start here.
WOW! A very good read, direct and to the point. Thanks a lot, not only for myself but for the many others who will, I hope, read it, recognize its clarity of voice and vision, and come to the only possible conclusion about the Starchild, based on the evidence.
Gary M.
There are two basic forms of plants and animals: wild and domesticated. The wild ones far outnumber the domesticated ones, which may explain why vastly more research is done on the wild forms. But it could just as easily be that scientists shy away from the domesticated ones because the things they find when examining them are so far outside the accepted evolutionary paradigm.
Nearly all domesticated plants are believed to have appeared between 10,000 and 5,000 years ago, with different groups coming to different parts of the world at different times. Initially, in the so-called Fertile Crescent of modern Iraq, Syria and Lebanon, came wheat, barley and legumes, among other varieties. Later on, in the Far East, came wheat, millet, rice and yams. Later still, in the New World, came maize (corn), peppers, beans, squash, tomatoes and potatoes.
Many have "wild" predecessors that were apparently a starting point for the domesticated variety, but others — like many common vegetables — have no obvious precursors. But for those that do, such as wild grasses, grains and cereals, how they turned into wheat, barley, millet, rice, etc. is a profound mystery.
No botanist can conclusively explain how wild plants gave rise to domesticated ones. The emphasis here is on "conclusively". Botanists have no trouble hypothesizing elaborate scenarios in which Neolithic (New Stone Age) farmers somehow figured out how to hybridize wild grasses, grains and cereals, not unlike Gregor Mendel when he cross-bred pea plants to figure out the mechanics of genetic inheritance. It all sounds so simple and so logical, almost no one outside scientific circles ever examines it closely.
Gregor Mendel never bred his pea plants to be anything other than pea plants. He created short ones, tall ones and different-colored ones, but they were always pea plants that produced peas. (Pea plants are a domesticated species, too, but that is irrelevant to the point to be made here.)
On the other hand, those New Stone Age farmers who were fresh out of their caves and only just beginning to turn soil for the first time (as the "official" scenario goes), somehow managed to transform the wild grasses, grains and cereals growing around them into their domesticated "cousins". Is that possible? Only through a course in miracles!
Actually, it requires countless miracles within two large categories of miracles. The first was that the wild grasses and grains and cereals were useless to humans. The seeds and grains were maddeningly small, like pepper flakes or salt crystals, which put them beyond the grasping and handling capacity of human fingers. They were also hard, like tiny nutshells, making it impossible to convert them to anything edible. Lastly, their chemistry was suited to nourishing animals, not humans.
So wild varieties were entirely too small, entirely too tough and nutritionally inappropriate for humans. They needed to be greatly expanded in size, greatly softened in texture and overhauled at the molecular level — which would be an imposing challenge for modern botanists, much less Neolithic farmers.
Despite the seeming impossibility of meeting those daunting objectives, modern botanists are confident the first sodbusters had all they needed to do it: time and patience. Over hundreds of generations of selective crossbreeding, they consciously directed the genetic transformation of the few dozen that would turn out to be most useful to humans.
And how did they do it? By the astounding feat of doubling, tripling and quadrupling the number of chromosomes in the wild varieties! In a few cases, they did better than that. Domestic wheat and oats were elevated from an ancestor with seven chromosomes to their current 42 — an expansion by a factor of six. Sugar cane was expanded from a 10-chromosome ancestor to the 80-chromosome monster it is today — a factor of eight.
The chromosomes of others, like bananas and apples, were only multiplied by factors of two or three, while peanuts, potatoes, tobacco and cotton, among others, were expanded by factors of four. This is not as astounding as it sounds, because many wild flowering plants and trees have multiple chromosome sets.
But that brings up what Charles Darwin himself called the "abominable mystery" of flowering plants. The first ones appear in the fossil record between 150 and 130 million years ago, primed to multiply into over 200,000 known species. But no one can explain their presence because there is no connective link to any form of plants that preceded them.
It is as if, dare I say it?, they were brought to Earth by something akin to You Know What. If so, then it could well be that they were delivered with a built-in capacity to develop multiple chromosome sets, and somehow our Neolithic forebears cracked the codes for the ones most advantageous to humans.
However the codes were cracked, the great expansion of genetic material in each cell of the domestic varieties caused them to grow much larger than their wild ancestors. As they grew, their seeds and grains became large enough to be easily seen and picked up and manipulated by human fingers. Simultaneously, the seeds and grains softened to a degree where they could be milled, cooked and consumed. And at the same time, their cellular chemistry was altered enough to begin providing nourishment to humans who ate them. The only word that remotely equates with that achievement is: miracle.
Of course, "miracle" implies that there was actually a chance that such complex manipulations of nature could be carried out by primitive yeomen in eight geographical areas over 5,000 years. This strains credulity because, in each case, in each area, someone actually had to look at a wild progenitor and imagine what it could become, or should become, or would become.
Then they somehow had to ensure that their vision would be carried forward through countless generations that had to remain committed to planting, harvesting, culling and crossbreeding wild plants that put no food on their tables during their lifetimes, but which might feed their descendants in some remotely distant future.
It is difficult to try to concoct a more unlikely, more absurd, scenario, yet to modern-day botanists it is a gospel they believe with a fervor that puts many "six day" Creationists to shame. Why?
Because to confront its towering absurdity would force them to turn to You Know What for a more logical and plausible explanation.
To domesticate a wild plant without using artificial (i.e., genetic) manipulation, it must be modified by directed crossbreeding, which is only possible through the efforts of humans. So the equation is simple.
-
- Firstly, wild ancestors for many (but not all) domestic plants do seem apparent.
- Secondly, most domesticated versions did appear from 10,000 to 5,000 years ago.
- Thirdly, the humans alive at that time were primitive barbarians.
- Fourthly, in the past 5,000 years, no plants have been domesticated that are nearly as valuable as the dozens that were "created" by the earliest farmers all around the world.
Put an equal sign after those four factors and it definitely does not add up to any kind of Darwinian model.
Botanists know they have a serious problem here, but all they can suggest is that it simply had to have occurred by natural means because no other intervention — by God or You Know What — can be considered under any circumstances.
That unwavering stance is maintained by all scientists, not just botanists, to exclude overwhelming evidence such as the fact that in 1837 the Botanical Garden in St Petersburg, Russia, began concerted attempts to cultivate wild rye into a new form of domestication. They are still trying, because their rye has lost none of its wild traits, especially the fragility of its stalk and its small grain. Therein lies the most embarrassing conundrum botanists face.
To domesticate a wild grass like rye or any wild grain or cereal (which was done time and again by our Neolithic forebears), two imposing hurdles must be cleared. These are the problems of "rachises" and "glumes", which I discuss in my book, Everything You Know Is Wrong ; Book One: Human Origins (pp. 283-285) (Adamu Press, 1998). Glumes are botany's name for husks, the thin covers of seeds and grains that must be removed before humans can digest them. Rachises are the tiny stems that attach seeds and grains to their stalks.
While growing, glumes and rachises are strong and durable, so rain won't knock the seeds and grains off their stalks. At maturity, they become so brittle that a breeze will shatter them and release their cargo to propagate. Such a high degree of brittleness makes it impossible to harvest wild plants because every grain or seed would be knocked loose during the harvesting process.
So, in addition to enlarging, softening and nutritionally altering the seeds and grains of dozens of wild plants, the earliest farmers also had to figure out how to finely adjust the brittleness of every plant's glumes and rachises.
That adjustment was of extremely daunting complexity, perhaps more complex than the transformational process itself. The rachises had to be toughened enough to hold seeds and grains to their stalks during harvesting, yet remain brittle enough to be collected easily by human effort during what has come to be known as "threshing".
Likewise, the glumes had to be made tough enough to withstand harvesting after full ripeness was achieved, yet still be brittle enough to shatter during the threshing process. And — here's the kicker — each wild plant's glumes and rachises required completely different degrees of adjustment, and the final amount of each adjustment had to be perfectly precise! In short, there is not a snowball's chance that this happened as botanists claim it did.
As with plants, animal domestication followed a pattern of development that extended 10,000 to 5,000 years ago. It also started in the Fertile Crescent, with the "big four" of cattle, sheep, goats and pigs, among other animals. Later, in the Far East, came ducks, chickens and water buffalo, among others. Later still, in the New World, came llamas and vicuna. This process was not simplified by expanding the number of chromosomes.
All animals — wild and domesticated — are diploid, which means they have two sets of chromosomes, one from each parent. The number of chromosomes varies as widely as in plants (humans have 46), but there are always only two sets (humans have 23 in each).
The only "tools" available to Neolithic herdsmen were those available to farming kinsmen: time and patience. By the same crossbreeding techniques apparently utilized by farmers, wild animals were selectively bred for generation after generation until enough gradual modifications accumulated to create domesticated versions of wild ancestors. As with plants, this process required anywhere from hundreds to thousands of years in each case, and was also accomplished dozens of times in widely separated areas around the globe.
Once again, we face the problem of trying to imagine those first herdsmen with enough vision to imagine a "final model", to start the breeding process during their own lifetimes and to have it carried out over centuries until the final model was achieved. This was much trickier than simply figuring out which animals had a strong pack or herding instinct that would eventually allow humans to take over as "leaders" of the herd or pack.
For example, it took unbridled courage to decide to bring a wolf cub into a campsite with the intention of teaching it to kill and eat selectively and to earn its keep by barking at intruders (adult wolves rarely bark). And who could look at the massive, fearsome, ill-tempered aurochs and visualize a much smaller, much more amiable cow? Even if somebody could have visualized it, how could they have hoped to accomplish it? An aurochs calf (or a wolf cub, for that matter), carefully and lovingly raised by human "parents", would still grow up to be a full-bodied adult with hardwired adult instincts.
However it was done, it wasn't by crossbreeding. Entire suites of genes must be modified to change the physical characteristics of animals. (In an interesting counterpoint to wild and domesticated plants, domesticated animals are usually smaller than their wild progenitors.) But with animals, something more, something ineffable, must be changed to alter their basic natures from wild to docile. To accomplish it remains beyond modern abilities, so attributing such capacity to Neolithic humans is an insult to our intelligence.
All examples of plant and animal "domestication" are incredible in their own right, but perhaps the most incredible is the cheetah. There is no question it was one of the first tamed animals, with a history stretching back to early Egypt, India and China. As with all such examples, it could only have been created through selective breeding by Neolithic hunters, gatherers or early farmers. One of those three must get the credit.
The cheetah is the most easily tamed and trained of all the big cats. No reports are on record of a cheetah killing a human. It seems specifically created for high speeds, with an aerodynamically designed head and body. Its skeleton is lighter than other big cats; its legs are long and slim, like the legs of a greyhound. Its heart, lungs, kidneys and nasal passages are enlarged, allowing its breathing rate to jump from 60 per minute at rest to 150 bpm during a chase. Its top speed is 70 miles per hour, while a thoroughbred tops out at around 38 mph. Nothing on a savanna can outrun it. It can be outlasted, but not outrun.
Cheetahs are unique because they combine physical traits of two distinctly different animal families: dogs and cats. They belong to the family of cats, but they look like long-legged dogs. They sit and hunt like dogs. They can only partially retract their claws, like dogs instead of cats.
Their paw pads are thick and hard like a dog's, but to climb trees they use the first claw on their front paws in the same way a cat does. The light-colored fur on their body is like the fur of a short-haired dog, but the black spots on their bodies are inexplicably the texture of cat's fur. They contract diseases that only dogs suffer from, but they also get "cat only" diseases.
There is something even more inexplicable about cheetahs. Genetic tests have been done on them, and the surprising result was that in the 50 specimens tested they were all, every one, genetically identical with each other! This means the skin or internal organs of any of the thousands of cheetahs in the world could be switched with the organs of any other cheetah and not be rejected. The only other place such physical homogeneity is seen is in rats and other animals that have been genetically altered in laboratories.
Cheetahs stand apart, of course, but all domesticated animals have traits that are not explainable in terms that stand up to rigorous scientific scrutiny. Rather than deal with the embarrassment of confronting such issues, scientists studiously ignore them and, as with the mysteries of domesticated plants, explain them away as best they can. For the cheetah, they insist it simply cannot be some kind of weird genetic hybrid between cats and dogs, even though the evidence points squarely in that direction. And why? Because that, too, would move cheetahs into the forbidden zone occupied by You Know What.
The problem of the cheetahs' genetic uniformity is explained by something now known as the "bottleneck effect". What it presumes is that the wild cheetah population — which must have been as genetically diverse as its long history indicates — at some recent point in time went into a very steep population decline that left only a few breeding pairs alive. From that decimation until now, they have all shared the same restricted gene pool.
Unfortunately, there is no record of any extinction events that would selectively remove cheetahs and leave every other big cat to develop its expected genetic variation. So, as unlikely as it seems, the "bottleneck" theory is accepted as another scientific gospel.
Here it is appropriate to remind scientists of Carl Sagan's famous riposte when dealing with their reviled pseudoscience: "Extraordinary claims require extraordinary evidence." It seems apparent that Sagan learned that process in-house.
It also leads us, finally, to a discussion of humans, who are so genetically recent that we, too, have been forced into one of those "bottleneck effects" that attempt to explain away the cheetah.
Like all plants and animals whether wild or domesticated, humans are supposed to be the products of slight, gradual improvements to countless generations spawned by vastly more primitive forebears. This was firmly believed by most scientists in the 1980s, when a group of geneticists decided to try to establish a more accurate date for when humans and chimpanzees split from their presumed common ancestor.
Paleontologists used fossilized bones to establish a timeline that indicated the split came between five and eight million years ago. That wide bracket could be narrowed, geneticists believed, by charting mutations in human mitochondrial DNA — small bits of DNA floating outside the nuclei of our cells. So they went to work collecting samples from all over the world.
When the results were in, none of the geneticists could believe it. They had to run their samples through again and again to be certain. Even then, there was hesitancy about announcing it. Everyone knew there would be a firestorm of controversy, starting with the paleontologists — who would be given the intellectual equivalent of a black eye and a bloody nose and their heads dunked into a toilet for good measure! This would publicly embarrass them in a way that had not happened since the Piltdown hoax was exposed.
Despite the usual scientific practice of keeping a lid on data that radically differs from a current paradigm, the importance of this new evidence finally outweighed concern for the image and feelings of paleontologists. The geneticists gathered their courage and stepped into the line of fire, announcing that humans were not anywhere near the official age range of eight to five million years old. Humans were only about200,000 years old. As expected, the howls of protest were deafening.
Time and much more testing of mitochondrial DNA and male Y-chromosomes now make it beyond doubt that the geneticists were correct. And the paleontologists have come to accept it because geneticists were able to squeeze humans through the same kind of "bottleneck effect" they used to try to ameliorate the mystery of cheetahs.
By doing so, they left paleontologists still able to insist that humans evolved from primitive forebears walking upright on the savannas of Africa as long ago as five million years, but that between 100,000 and 200,000 years ago "something" happened to destroy nearly all humans alive at the time, forcing them to reproduce from a small population of survivors.
That this "something" remains wholly unknown is a given, although Creationists wildly wave their hands like know-it-alls at the back of a classroom, desperate to suggest it was the Great Flood. But because they refuse to move away from the biblical timeline of the event (in the range of 6,000 years ago), nobody can take them seriously. Still, it seems the two sides might work together productively on this crucial issue. If only...
Apart from disputes about the date and circumstances of our origin as a species, there are plenty of other problems with humans. Like domesticated plants and animals, humans stand well outside the classic Darwinian paradigm. Darwin himself made the observation that humans were surprisingly like domesticated animals. In fact, we are so unusual relative to other primates that it can be solidly argued that we do not belong on Earth at all, that we are not even from Earth, because we do not seem to have developed here.
We are taught that, by every scientific measure, humans are primates very closely related to all other primates, especially chimpanzees and gorillas. This is so ingrained in our psyches that it seems futile even to examine it, much less to challenge it. But we will.
-
Bones. Human bones are much lighter than comparable primate bones. For that matter, our bones are much lighter than the bones of every "pre-human" ancestor through to Neanderthal. The ancestor bones look like primate bones; modern human bones do not.
-
Muscle. Human muscles are significantly weaker than comparable muscles in primates. Pound for pound, we are five to ten times weaker than any other primate. Any pet monkey is evidence of that. Somehow, getting "better" made us much, much weaker.
-
Skin. Human skin is not well adapted to the amount of sunlight striking Earth. It can be modified to survive extended exposure by greatly increasing melanin (its dark pigment) at its surface, which only the black race has achieved. All others must cover themselves with clothing or frequent shade or both, or sicken from radiation poisoning.
-
Body Hair. Primates need not worry about direct exposure to sunlight because they are covered from head to toe in a distinctive pattern of long body-hair. Because they are quadrupeds (move on all fours), the thickest hair is on their back, the thinnest on the chest and abdomen. Humans have lost the all-over pelt, and we have completely switched our area of thickness to the chest and abdomen while wearing the thin part on our back.
-
Fat. Humans have ten times as many fat cells attached to the underside of their skin as primates. If a primate is wounded by a gash or tear in the skin, when the bleeding stops the wound's edges lie flat near each other and can quickly close the wound by a process called "contracture". In humans, the fat layer is so thick that it pushes up through wounds and makes contracture difficult if not impossible. Also, contrary to the propaganda to try to explain this oddity, the fat under human skin does not compensate for the body hair we have lost. Only in water is its insulating capacity useful; in air, it is minimal at best.
-
Head Hair. All primates have head hair that grows to a certain length and then stops. Human head hair grows to such lengths that it could be dangerous in a primitive situation. Thus, we have been forced to cut our head hair since we became a species, which may account for some of the sharp flakes of stones that are considered primitive hominid "tools".
-
Fingernails and Toenails. All primates have fingernails and toenails that grow to a certain length and then stop, never needing paring. Human fingernails and toenails have always needed paring. Again, maybe those stone "tools" were not only for butchering animals.
-
Skulls. The human skull is nothing like the primate skull. There is hardly any fair morphological comparison to be made, apart from the general parts being the same. Their design and assembly are so radically different as to make attempts at comparison useless.
-
Brains. The comparison here is even more radical because human brains are so vastly different. (To say "improved" or "superior" is unfair and not germane, because primate brains work perfectly well for what primates have to do to live and reproduce.)
-
Locomotion. The comparison here is easily as wide as the comparison of brains and skulls. Humans are bipedal; primates are quadrupeds. That says more than enough.
-
Speech. Human throats are completely redesigned relative to primate throats. The larynx has dropped to a much lower position, so humans can break typical primate sounds into the tiny pieces of sound (by modulation) that have come to be human speech.
-
Sex. Primate females have oestrous cycles and are sexually receptive only at special times. Human females have no oestrous cycle in the primate sense. They are continually receptive to sex. (Unless, of course, they have the proverbial headache!)
-
Chromosomes. This is the most inexplicable difference of all. Primates have 48 chromosomes. Humans are considered vastly superior to them in a wide array of areas, yet somehow we have only 46 chromosomes! This begs the question of how we could lose two full chromosomes — which represents a lot of DNA — in the first place, and in the process become so much better. Nothing about it makes logical sense.
-
Genetic Disorders. As with all wild animals (plants, too), primates have relatively few genetic disorders spread throughout their gene pools. Albinism is one that is common to many animal groups as well as humans. But albinism does not stop an animal with it from growing up and passing the gene for it into the gene pool. Mostly, though, serious defects are quickly weeded out in the wild. Often, parents or others in a group will do the job swiftly and surely, so wild gene pools stay relatively clear. In contrast, humans have over 4,000 genetic disorders, and several of those will absolutely kill every victim before reproduction is possible. This begs the question of how such defects could possibly get into the human gene pool in the first place, much less how they remain so widespread.
-
Genetic Relatedness. A favorite Darwinist statistic is that the total genome (all the DNA) of humans differs from chimpanzees by only 1% and from gorillas by 2%. This makes it seem as if evolution is indeed correct and that humans and primates are virtually kissing cousins. However, what they don't stress is that 1% of the human genome's three billion base pairs is 30 million base pairs — and to any You Know What that can adroitly manipulate genes, 30 million base pairs can easily add up to a tremendous amount of difference.
-
Everything Else. The above are the larger categories at issue in the discrepancies between primates and humans. There are dozens more listed as sub-categories below one or more of these.
To delve deeper into these fascinating mysteries, check The Scars of Evolution by Elaine Morgan (Oxford University Press, 1990). Her work is remarkable. And for a more in-depth discussion of the mysteries within our genes and those of domesticated plants and animals, see Everything You Know Is Wrong.
When all of the above is taken together — the inexplicable puzzles presented by domesticated plants, domesticated animals and humans — it is clear that Darwin cannot explain it, modern scientists cannot explain it, not Creationists nor Intelligent Design proponents. None of them can explain it, because it is not explainable in only Earthbound terms.
We will not answer these questions with any degree of satisfaction until our scientists open their minds and squelch their egos enough to acknowledge that they do not, in fact, know much about their own backyard. Until that happens, the truth will remain obscured.
My personal opinion, which is based on a great deal of independent research in a wide range of disciplines relating to human origins, is that ultimately Charles Darwin will be best known for his observation that humans are essentially like domesticated animals.
I believe that what Darwin observed with his own eyes and research is the truth, and that modern scientists would see it as clearly as he did if only they had the motivation or the courage to seek it out. But for now, they don't, so, until then, we can only poke and prod at them in the hope of some day getting them to notice our complaints and address them. In order to poke and prod successfully, more people have to be alerted to the fact that another scientific fraud is being perpetrated.
Future editions of Icons of Evolution will discuss the current era when scientists ridiculed, ignored or simply refused to deal with a small mountain of direct, compelling evidence that outside intervention has clearly been at work in the genes of domesticated plants, animals and humans. You Know What has left traces of their handiwork all over our bodies, all through our gene pools. All that will be required for the truth to come out is for a few "insiders" to break ranks with their brainwashed peers.
Look to the younger generation. Without mortgages to pay, families to raise and retirements to prepare for, they can find the courage to act on strong convictions. Don't expect it of anyone over forty, possibly even thirty. But somewhere in the world, the men and women have been born who will take Darwinism down and replace it with the truth.
I will end this eBook with a remarkable letter sent to me by email on February 12, 1999, from a man who, to protect himself from vindictive peers, called himself “DNA Deep Throat.”
He is a highly skilled geneticist whom I have since come to know by name, though he still must stay anonymous because his mainstream geneticist peers remain as vindictive as ever.
This man was instrumental in the early days of my dealings with the now well-known Starchild Skull, but in 1999, I was just getting started and in much need of help to figure out what it was [For more understanding of the Starchild Skull, a book about it, The Starchild Skull, is available at www.StarchildProject.com, and also through www.LloydPye.com, as is an eBook about it.]
An early theory about the Starchild was that it might be a human-alien hybrid, and a question that kept being asked was, “How can a human and an alien hybridize? Won’t they be too far apart genetically? How could that happen?”
As we have seen, that question is essentially the same one we have to answer in regard to Zana. With both Zana and the Starchild in mind, read the words sent to me in 1999, and you will have the same degree of insight they imparted to me.
Dear Mr. Pye:
I agree with your conclusions and will give you a few hints, if you wish, (speaking) as a “DNA Deep Throat.” First, look up the huge discontinuities between humans and the various apes for: (1) Whole mitochondrial DNA; (2) genes for the Rh Factor; (3) and human Y chromosomes, among others.
Regarding #3, I refer you to K.D. Smith’s 1987 study titled “Repeated DNA sequences of the human Y chromosome.” It says “Most human Y chromosome sequences thus far examined do not have homologues [same relative position or structure] on the Y chromosomes of other primates.”
Human female X chromosomes do look somewhat apelike, but not the male’s Y. This means that if humans are a crossbred species, the cross had to be between a female ape-like creature (i.e, “creature of Earth”) and a male being from elsewhere.
What the evolutionists do is find certain genes which look very similar between man and ape, then they make a “tree of descent” while ignoring those huge impassable abysses of difference elsewhere.
Also, by certain methods of DNA dating, one can tell that numerous genes have been recently added to the human genome. If workers in my field were to say such things openly, we would be ostracized and forced to live in a tent. Any work along these lines would be rejected without any form of appeal. So what can we do?
Sincerely,
DNA Deep Throat
Let’s leave it there, with DNA Deep Throat’s parting question: What can we do when we are up against a monolith like mainstream science, which won’t forsake its cherished dogmas until those dogmas are crammed down their throats?
The End